Logic of thinking. Part 5. Brain waves
So, we have come to the description of one of the key principles of the described model.
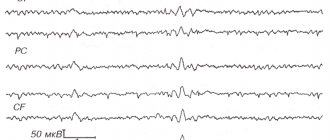
This principle has not previously been used either in neural networks or in describing the functioning of the brain. In this regard, I highly recommend that you read the previous parts. At a minimum, you need to read the fourth part, without which what is described below will be completely incomprehensible. In the previous part, we talked about the fact that neuronal activity is divided into evoked and background. Echoes of background activity are observed by taking an electroencephalogram. The recorded signals have a complex shape and depend on the location of application of the electrodes to the head, but, nevertheless, individual harmonic components can be clearly traced in them. The main rhythms are named:
- alpha rhythm (from 8 to 13 Hz);
- beta rhythm (15 to 35 Hz);
- gamma rhythm (from 35 to 100 Hz);
- delta rhythm (from 0.5 to 4 Hz);
- theta rhythm (from 5 to 7 Hz);
- sigma-ri (13 to 14 Hz).
The nature of rhythms is traditionally correlated with the oscillatory properties of neurons. Thus, the behavior of a single neuron is described by the Hodgkin-Huxley equations (Hodgkin, 1952):
These equations determine the autowave process, that is, they instruct the neuron to generate impulses. The Hodgkin-Huxley model is difficult to model. Therefore, there are quite a lot of its simplifications that preserve the basic generating properties. The most popular models: Fitzhugh-Nagumo (Fitzhugh, 1961), Morris-Lecar (Morris C., Lecar H., 1981), Hindmarsh-Rose (Hindmarsh JL, and Rose RM, 1984). Many models, for example, Hindmarsh-Rose, allow you to simulate both burst activity and isolated spikes (figure below).
Burst activity and spikes occurring in a Hindmarsh-Rose neuron model
By combining neurons that generate their own impulses into structures that resemble the structure of the real cortex, it is possible to reproduce various effects characteristic of the group activity of real neurons. For example, one can achieve global synchronization of neural activity or cause the appearance of waves. The most famous models are Wilson-Cowan (HR Wilson and JD Cowan, 1972) and Kuromoto (Kuramoto, 1984).
The electroencephalogram records echoes of the joint activity of neurons, but it is clear that this activity has a certain spatiotemporal organization. Methods for optical observation of cortical activity make it possible to see it live. A section of the cortex of the experimental animal is exposed and a special dye is injected that is sensitive to changes in electrical potential. Under the influence of total fluctuations in the membrane potential of neurons, such a dye changes its spectral properties. And although these changes are extremely small, they can nevertheless be recorded, for example, using a diode array that acts as a high-speed video camera. Optical methods do not allow one to look deep into the cortex and trace the activity of individual neurons, but they make it possible to get a general idea of the flow of wave processes on its surface.
Scheme of optical recording of cortical activity (Michael T. Lippert, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu, 2007)
It turned out that the rhythms of the brain correspond to waves that arise in point sources and spread across the cortex, like circles on water. True, the wave front on water diverges strictly in a circle, but the wave front of the activity of brain neurons can propagate in a more complex way. The figure below shows patterns of wave propagation over a 5-millimeter section of the rat's cerebral cortex.
A picture of the propagation of a wave of activity in a section of the rat’s cerebral cortex. The potential is shown as a gradient from blue to red. 14 frames at 6 millisecond intervals cover one wave propagation cycle (84 milliseconds - 12 Hz) (Michael T. Lippert, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu, 2007)
Very interesting and revealing video recordings of wave activity are presented in (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y. Wu, 2007). It is shown there that waves can become denser, reaching the boundary of a crustal zone, can be reflected from another zone and create a counter wave, can propagate in double spirals and create vortices.
Wave compression and reflection in primary visual cortex (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y. Wu, 2007)
Below is a video from the same work (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y. Wu, 2007):
Using models of oscillating neurons, it is possible to obtain similar wave patterns in computer experiments. But such a pulsation in itself does not make much sense. It is quite reasonable to assume that waves of activity are a mechanism for transferring and processing information. But attempts in traditional models to explain the nature of this mechanism have not yielded tangible results. It seems logical to assume that information is encoded by the frequency and phase characteristics of neural signals. But this leads to the need to take into account interference processes and raises more questions than it brings clarity.
There is a hypothesis that the waves “scan” the cortex, reading information from it for its subsequent transmission (Pitts W., McCulloch WS, 1947). This seems reasonable enough, at least as far as alpha waves and the primary visual cortex are concerned. The authors of this hypothesis, McCulloch and Peets, emphasized that scanning helps explain one important phenomenon. The fact is that information from the primary visual cortex is projected further along the fibers of the white matter through bundles of axons, which are clearly insufficient in volume to simultaneously transmit the entire state of this cortical zone. This means, they concluded, scanning allows the use of not only spatial, but also temporal code, which ensures sequential transmission of the required information.
This problem of narrowness of the projecting bundles exists not only for the primary visual cortex, but also for all other areas of the brain. The number of fibers in the projection pathways is much less than the number of neurons that form the spatial pattern of activity. The connections between zones are clearly not able to transmit the entire spatially distributed signal in parallel, which means that an understanding of how the information transmitted through them is compressed and decompressed occurs. The scanning assumption, although it does not answer the question about the encoding mechanism itself, nevertheless allows us to pose the right questions.
Our model in explaining brain rhythms is based on the above description of the nature of metabotropic activity. Using the idea that not only neuronal synapses, but also metabotropic receptive clusters are involved in the process of creating rhythms, allows us to obtain a result that is qualitatively different from all classical theories. But before I get into this, I want to give a warning. I will deliberately describe simplified idealized models, matching them with certain processes characteristic of the real brain, but without claiming that the brain works exactly like that. Our task is to show the basic principles, understanding that evolution has gone far ahead and their true implementation is much more cunning. An analogy can be drawn with the development of computer technology. A modern computer is quite complex, and if we begin to describe the basic principles of classical computing devices, it turns out that in their pure form they are almost impossible to find in modern systems. Basic representation - the processor reads the program and data from memory, performs the actions prescribed by the program on the data, and writes the results back to memory. Now add to this the use of caches of various levels, multithreading, hyperthreading, parallel computing using local, group and shared memory, and the like. And it turns out that it is difficult to find literal compliance with simple rules in a real computer. Actually, all this must be taken into account when comparing the subsequent description with the work of a real brain.
So, let's take a model of a cortical area on which we will create a compact pattern of evoked activity. For now, let’s leave the question of how this pattern arose. Let's just assume that there are elements that have a constant pulse signal. In the figure below, neurons that form this pattern are marked in red. In real cortex, this corresponds to axons that transmit burst activity, that is, they produce a series of spikes at a high frequency. These axons may belong to neurons in the same cortical area that are in a state of evoked excitation, or they may be projection fibers coming from other parts of the brain.
Pattern of evoked activity
Now let's force free neurons to generate rare random impulses. In this case, we impose the condition that a random spike requires a certain level of surrounding activity. This means that random spikes can only occur in the vicinity of already active neurons. In our case, they will appear in the vicinity of the evoked activity pattern. In the figure below, random spikes are shown in green.
Typically, all neuronal activity that is not evoked is called spontaneous or background. This is not very successful, as it sets one up to perceive all this activity as random. Next we will show that, in general, background activity is strictly predetermined and not at all random. And only a small part of it is truly completely random spikes. We have now created such random spikes around the active pattern.
First step of modeling. Spontaneous activity versus evoked activity
Let's model metabotropic receptive clusters using our formal neurons. To do this, we will give neurons the opportunity to remember, when necessary, the picture of activity in the immediate environment. Moreover, we will not limit ourselves to one picture for one neuron, as it would be if we memorized at synapses, but will allow each neuron to store many such pictures.
Let’s make sure that the presence of our own random spike and high surrounding activity will be the basis each time to remember the pattern of surrounding impulses (figure below). Next, our neurons will produce single spikes every time one of these memorized local patterns is repeated. In order not to confuse them with random spikes, we will call these spikes wave spikes.
The activity recording area for one of the neurons that produced a spontaneous spike (it is in the center of the square). When modeling, for simplicity, square receptive fields were used, which, although it does not correspond to the tracking fields of real neurons, does not qualitatively affect the result
As a result, at the next modeling step (figure below), we will get a picture in which there will be the same as before, neurons with evoked activity (red), neurons that responded to local images memorized at the previous step (white), and neurons that generated spontaneous spike (green).
Second modeling step. Red – evoked activity, white – wave activity, green – spontaneous activity
By repeating the modeling steps, we obtain cortical propagating activity with a specific unique randomly generated pattern. Now we introduce the condition of neuron fatigue. Let's make it so that after several metabotropic (wave) spikes, neurons will lose the ability to generate new impulses for the time required for relaxation. This will lead to the fact that the activity will not spread in a continuous area, but in pulsating waves scattering from their source.
To prevent “clogging,” let’s block spontaneous activity in areas where there is a large number of relaxing neurons. As a result, we will get a ring of neurons with wave activity, running away from the pattern of evoked activity (figure below).
Wave front propagation. Blue – neurons in a state of relaxation
The resulting wave, as it moves away from its center, will lose front density and sooner or later will die out. In the figure below you can trace the first cycle of her life.
First cycle of ID wave propagation
After the relaxation period has passed, a new wave will start. Now it will be picked up by neurons that have already undergone training and will spread a little further. With each new cycle, the wave will expand the boundaries of its propagation until it reaches the edges of the zone. To prevent an unlimited increase in the density of the wave front, we will introduce one more restriction: we will prohibit spontaneous activity for neurons when the total activity around them exceeds a certain threshold.
After some time, we will receive a cortex trained to propagate a wave of activity that is unique in its pattern, corresponding to the initially specified pattern of evoked activity (figure below).
Wave propagation over the already trained cortex (after 200 training cycles)
Below is a video of the wave learning process:
Please note: when the cortex has already been trained to propagate a wave from a certain pattern, spontaneous (random) spikes disappear. Such an “experienced” wave is formed due to wave, non-random spikes. Random spikes appear only during training, when the wave front reaches untrained or poorly trained areas. At this moment, spontaneous impulses create a random continuation of the wave pattern, which is immediately remembered on the surface of the neurons participating in this wave. But as soon as the training ends, the wave from the pattern familiar to the cortex begins to spread along the already created wave pattern, each beat exactly repeating the same non-random pattern along its path.
Training the cortex does not have to be gradual. When modeling by selecting parameters, it is possible to ensure that the generated wave is self-sufficient. That is, so that it does not fade out in the first bars, but immediately spreads to the entire space (picture below).
An example of a training wave propagation that does not decay in the first cycle
Since the width of the wave front is constant, its area increases with distance from the source. This corresponds to more neurons being involved in the propagation of activity. If we trace the total potential created by them, we will get graphs that resemble what we see on encephalograms (figure below).
Single source model rhythmic activity graph
Please note that the appearance of the rhythms observed in such “encephalograms” is not a ri of the cortex. That is, not synchronization of joint bursts of activity, as expected in most existing models, but a change in the number of neurons involved in the propagation of divergent waves. If new waves are emitted before the fronts of the previous waves disappear, then their addition will give a smoother picture. With a certain selection of parameters, the model reproduces situations where the propagation of waves is practically not accompanied by total rhythmic activity. This can be correlated with the fact that about 10 percent of people do not show wave activity on electroencephalograms.
If we now take a new pattern of evoked activity, the cortex will create waves that spread out from it. Moreover, such a cortex learns to create waves for any stable pattern of evoked activity. Due to the fact that the same neuron can store many local images, it can be part of many different waves at once, corresponding to different patterns. If we want greater uniqueness of waves, then it is enough, as the neuron accumulates local images stored by it, to reduce the probability of its spontaneous spike.
For stored local images, it makes sense to introduce a consolidation mechanism. That is, not to fix the image immediately for centuries, but to set the time during which the image should be repeated a certain number of times. Since wave propagation learning and the formation of stable patterns of evoked activity are parallel processes, consolidation can make it possible to erase traces of unsuccessful learning.
But the most important thing in all this is the uniqueness of each of the resulting wave stories. In any place through which a wave passes, it creates a unique pattern that is characteristic only of it. This means that if there is a pattern of evoked activity on the surface of the cortex that encodes an event, then the wave caused by it will spread information about this throughout the entire cortex. In each specific place, this wave will create its own unique pattern, unique only for this event. Any other event will create a different pattern in this place, associated with its wave. This means that, being in any place, we can tune in to “receive” a wave from any specific event and record when it occurs upon the arrival of a wave with the corresponding pattern.
In our model, it turns out that the fundamental properties of the crust are:
- the ability to generate waves diverging from patterns of evoked activity;
- the ability to learn, to conduct wave patterns that are unique and stable for each pattern;
- once trained, the ability to propagate waves of activity around stable patterns of evoked activity.
Each unique pattern of evoked activity corresponds to a wave that is unique in its pattern, which can be called the identifier of this pattern.
Now the learning process can be roughly represented as follows:
- statistically stable signals lead to the formation of detector neurons, which, due to synaptic plasticity, learn to respond to certain patterns;
- detector neurons related to the same pattern form a stable pattern;
- each stable pattern, as it is active, teaches the cortex to propagate a unique wave corresponding only to this pattern, which can be interpreted as the identifier of this pattern;
- Each place in the cortex where the corresponding waves propagate gets the opportunity to find out what the general information picture is at the current moment.
From the above, it follows that for its full-fledged operation, the brain must not only form neurons-detectors of various phenomena, but also train the cortex to propagate the corresponding information waves.
This fits well with the fact that rhythmic activity in children is weak immediately after birth and increases as they grow older. When we talk about brain structures that respond to certain phenomena, their obvious dualism arises in our concept. These structures are not described exclusively by any neurons or groups of neurons, they are also supplemented by identification waves that allow the dissemination of relevant information. But what’s interesting is that in each specific place through which the wave passes, it behaves in the same way as the pattern that gave rise to it. In an instantaneous time slice, it itself is a pattern that generates a continuation of the wave.
It is easy to see that such propagation of identification waves repeats the Huygens-Fresnel principle, which describes the propagation of light waves. Its essence is that each point of the light wave front can be considered as the center of a secondary disturbance. Each such center generates secondary spherical waves. The resulting light field is the interference of these waves. The envelope of the secondary waves becomes the wave front at the next moment in time, and so on.
Refraction (left) and interference (right) according to Huygens-Fresnel (Nordmann)
The difference in the nature of the pattern of evoked activity and the pattern that arises during the propagation of the wave can only be detected by analyzing the cause of the activity of the neurons included in them. For evoked activity, this is the pattern at the synapses; for the propagation pattern, this is the reaction of metabotropic receptors. That is, the brain’s reflection of any phenomena is both a pattern and a wave, and at any given moment in time, when we try to record the propagation of this wave, we will see a pattern that is different in design, but not in essence from what gave rise to the wave .
It is easy to see that the described dualism quite accurately corresponds to the dualism of elementary particles. As you know, every particle is both a particle and a wave. Depending on what circumstances we consider, a particle can exhibit both wave and corpuscular properties. Modern physics states this fact, but does not explain it. What we have described about the dualism of information images is, fortunately, somewhat clearer than the dualism of elementary particles. Of course, this begs the question, is it possible to extend these analogies and use them to understand the physical picture of the world? This is a very serious question, and later we will talk about whether such a coincidence is accidental or not.
Important consequences flow from the dualism of information images. Like particles with different spins, two patterns from different waves can be in the same spatial region without interfering with each other. Moreover, each of them causes the continuation of its own wave without introducing mutual distortions. If we create several patterns and train the cortex to propagate waves from each of them, then by activating these patterns together, we will be able to observe the passage of waves through each other while maintaining our own uniqueness (picture below).
Simulation of the passage of waves from two sources
This is very different from continuous wave processes. For example, when waves propagate in water or air, all molecules take part in the vibrations at once. The collision of two waves affects the entire contacting volume, which leads to interference. The behavior of each molecule is determined by the action of both waves at once. As a result, we get an interference pattern that is no longer as simple as each of the wave patterns separately. In our case, each of the waves propagates, affecting only a small part of all available neurons. When two information waves intersect, the neurons they have in common that can create interference are too few to influence the propagation of each of them. As a result, the waves retain their information picture, undistorted from meeting another wave.
A certain elegance of the described system also lies in the fact that neurons can simultaneously perform two functions. Each neuron can be a detector, tuning itself by changing synaptic weights to a specific image. Moreover, the same neuron, due to the information recorded on its metabotropic clusters, can take part in the propagation of various information waves.
It is worth feeling the difference between the properties of a neuron as a detector and as a participant in the propagation of wave activity. It would seem that in both cases he reacts to a certain picture of the activity of his environment. In reality, these are two fundamentally different properties. Relatively smooth adjustment of synaptic weights allows the neuron to learn to identify hidden factors. Instant fixation of images by extrasynaptic clusters ensures memorization of what is there right now. Synapses perceive a large-scale picture from the entire receptive field of the neuron. Metabotropic receptive clusters have access to a reduced pattern of activity, limited to the synapses that form their synaptic traps. An ordinary neuron has only one set of synaptic weights, but it can store tens and hundreds of thousands of identification images. Synaptic recognition produces evoked activity, that is, a long burst of impulses. Several detector neurons thereby create a pattern of evoked activity. Metabotropic recognition produces a single spike. The combination of such spikes creates the front of the identification wave.
During modeling, it is observed that the propagation range of the identifier depends on how active the wave-creating pattern is. By using the mechanism of consolidation, that is, by introducing criteria for forgetting that is triggered if the activity is not repeated properly, it is possible to obtain an effect when frequently repeating patterns train the entire cortex to propagate their waves, and rare combinations create areas of local propagation. Moreover, the sizes of these areas are not fixed, but willingly grow if the corresponding phenomenon begins to manifest itself more often. There is a certain reason for this behavior. It is possible that the real cortex behaves in a similar way.
When I managed to formulate the wave principles described above, modeling within the framework of our project moved to a new qualitative level. It felt like instead of a stone ax we suddenly had a jackhammer. In the following parts, I will show how the use of the wave neural network model allows us to explain many properties of the brain that previously seemed mysterious.
Continuation
If something is too brief, unclear or unclear, please write in the comments.
Previous parts: Part 1. Neuron Part 2. Factors Part 3. Perceptron, convolutional networks Part 4. Background activity
Beta rhythm
The beta rhythm is associated with higher cognitive processes and focusing attention. The frequency of Beta waves varies from 14 to 30 Hz, with an oscillation amplitude of up to 20 μV. The beta rhythm is recorded in the region of the anterior and central gyri and extends to the posterior central and frontal gyri.
The beta rhythm is the norm and is activated when a person is involved in the world around him, when observing ongoing processes and focusing on solving any current problems.
Beta waves are actively emitted in states of increased arousal, anxiety or stress, in addition, activation of Beta waves occurs during communication, for example, when talking.
When Beta waves are activated, blood pressure typically rises and metabolism increases dramatically.
The dominance of Beta waves is considered a state close to stress, regardless of the nature of a person’s emotions.
However, if the brain rarely functions in the Beta rhythm, the person experiences depression, poor memory and lack of normal concentration.
No special exercises are required to develop Beta rhythms.
It's enough to just live an active life. Any physical activity, communication, stress, fears, emotions help increase Beta rhythms.
Reading interesting books and caffeine also help increase beta waves.
Energy drinks and smoking also give a surge in beta wave activity, however, for a very short time and a temporary rise is certainly followed by a decline, as well as a depressed state caused by excessive energy consumption.
To reduce Beta rhythms, simply relax and give up negative thoughts. It should be noted that there are 3 types of Beta rhythms: low, medium and high.
Low Beta rhythms (13-16 hertz) - occur with weak involvement in the outside world. The left hemisphere of the brain is activated. There is no excitement, but the relaxed state also passes. With regular training at this frequency, you can develop the skill of managing attention and concentration.
Medium Beta rhythms (16-18 hertz) - occur with some involvement in the outside world. The right hemisphere of the brain is connected to work. There is some excitement. With regular training at this frequency, you can develop intellectual abilities and concentration skills.
High Beta rhythms (18-30 hertz) - occur with increased arousal, are characterized on the one hand by increased interest in the world around us, and on the other hand by severe stress and high blood pressure.
Gamma rhythm
The gamma rhythm is usually observed when solving problems that require maximum concentration. The frequency of the waves varies from 30 to 100 Hz, the amplitude usually does not exceed 15 μV. It is recorded in the precentral, frontal, temporal and parietal areas of the cerebral cortex. There are theories that disturbances in Gamma rhythms are one of the causes of schizophrenia. The brain oscillation frequency of 50 Hz is what some researchers of Buddhist practices call the state of Enlightenment. Also, it is precisely for this frequency of vibrations that the state of presence “here and now” is characteristic.
Delta rhythm
The delta rhythm (slow waves) occurs at very low levels of activity and is associated with restorative processes, especially during trance or deep dreamless sleep. Also, the Delta rhythm is characteristic of a state of very deep meditation - Dhyana (not relaxation, like the alpha rhythm).
The frequency of the waves varies from 0.1 to 4 Hz, the amplitude is 50–500 μV. The predominance of the Delta rhythm in a state of wakefulness leads to impairment of attention and other cognitive functions.
With a high amplitude of Delta waves, there is often increased intuition, as well as the ability to receive and process information on an unconscious level. This feature of Delta waves is actively used by healers and people with paranormal abilities when obtaining the necessary information.
The simplest and most effective way to achieve sustainable development of Delta waves is rhythmic breathing at a frequency of about 60 breaths per minute.