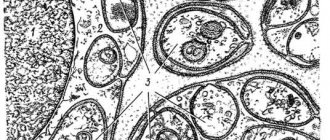
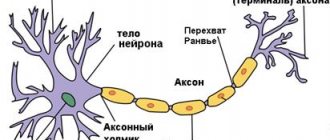
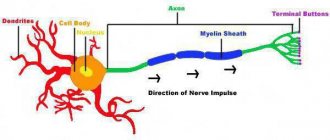
Строение
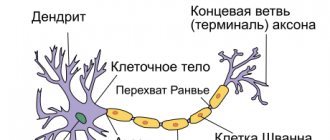
Общее строение нейрона выглядит следующим образом: есть тело (сома), в котором содержатся ядро и другие органоиды, и отростки — аксон и дендриты:
- Аксон
присутствует всего один — это отросток, по которому нервный импульс идет от данной клетки к другим. Другими словами, аксон — канал выхода сигнала. - Дендриты
, соответственно, — каналы входа сигналов, и их может быть как очень много, так и совсем мало. Количество дендритов зависит от типа нейрона, и об этом мы поговорим позже.
Рис. 1. Схема нейрона
Аксоны и дендриты
Аксоны — отростки, которые могут достигать в длину более метра. Чтобы сигнал не “рассеивался” по пути от одной клетки к другой, большинство аксонов в теле покрыты миелиновой оболочкой, состоящей из клеток нейроглии (общее обозначение вспомогательных клеток нервной ткани). Оболочка обеспечивает изоляцию одного аксона от других и не позволяет электрическому импульсу рассеяться. Благодаря миелиновой оболочке, проведение импульса по аксону осуществляется быстрее. Дендриты более короткие и не покрыты миелином.
Эндоплазматическая сеть (ЭПС) и комплекс Гольджи
Наиболее важными органоидами, помимо ядра, являются шероховатая ЭПС, имеющая рибосомы и осуществляющая синтез белков, и аппарат Гольджи, синтезирующий различные органические вещества и “упаковывающий” их в мембранные пузырьки. Почему эти системы так важны для функциональной деятельности нейрона — будет понятно далее.
Функциональная характеристика афферентных,эфферентных и вставочных нейронов.(понятие «нейрон»)
Нейроны – специализир кл-ки,способн приним,обрабатыв,кодир,хран,передав и воспроизв инф-ю,организовывать р-и на раздр-я,устанавл контакты с др.нейр и к-ми органов. Способн генерир электрич потенц и с их помощью перед инф-ю ч/з спец окончания – синапсы. Выполнению ф-й нейр способств нейромедиаторы,синтезир-ся в его аксоплазме. Разм 6-120мкм. В мозге-до 1011нейр. Для разл стр-р мозга характерно опред типы нейронной организации. Нейры образуют группы,ядра и т.п. Клеточные скопления образ серое в-во. Между ядрами и кл-ми проходят миелинове и безмиелинов волокна: аксоны и дендриты. Строение: функц выделяют след части: воспринимающую часть-дендриты,сома, на кот находится аксонный холмик; передающую чвсть-аксон. Сома: информ, трофическ по отношению к отросткам. Сод-ит:рибосомы,тигроидн в-во,КГ,лизосомы, пигменты-меланин и липофусцин, митохондрии, нейротруб,ядро с ядрышк. Дендриты- восприним импульсы от др нерв клеток. Аксонный холмик –генераторный пункт ПД.Аксон проводит возбуждение к др клеткам. Типы нейронов: по кол-ву отростков: псевдоуниполярные (2отростока,сливающ вблизи тела в1), биполярные(1аксон,1дендрит),мультиполярные(неск дендритов,1аксон).По направленности: афферентн, эфферентные и вставочные. От характера влияния: возбуждающие и тормозящие. От активности: фоново-активные(активны и без воздействия) имолчащие(активны в ответ на раздражение).От медиатора: адренергические, лолинергические, серотонинергические и др.
Афферентные нейроны.-нейр,воспринимающие инф-ю. как правило, имеют больш разветвлён.это хар-но для всех ур-ней ЦНС. В задн рогах спин мозга афер яв-ся чувствит нейроны малых размеров. Вставочные-(интернейроны)-обрабат инф-ю,получаемую от афер нейронов и передают их на другие вставочные или эффер н.,формируя нейр сети). Усиливают сигнал, удлиняет время сохр-я информации в центре. Всавочные нейроны могут быть возбуждающими(в новой коре,облегч передачу инф-и с одной группы нейр в другую); и тормозные(кот.возбуждаются прямыми сигналами или сигналами,идущими из того же центра по обратным связям). Эфферентные нейроны-нейр,передающие инф-ю от нервного центра к исполнительн орг или другим центрам НС. Связывают м/у собой разл стр-ры мозга,обеспечивая межполушарные и внутриполушарные связи,формируя функц сост-е мозга(утомление,обучение и проч).
Нейроглия.Её виды. Функциональная хар-ка и физиол. роль. Пульсация глиоцитов и ее значение.
Нейроглия – совокупность клет эл-тов нервной ткани, образованная спец клетками разл формы. Обнаружена она Вирховым. К-ки глии заполняют простр-ва м/у нейр,составляя 40%объёма мозга. Глиальные к-ки по разм в 3-4 р меньше нейр. Различают неск типов глии, в зав от сотава клеток: астроциты,олигодендроциты,микроглиоциты. Астроциты – многоотросчатые мелкие кл-ки с ядрами овальной формы и небольш числом хроматина. 7-25мкм. Расп. в сером в-ве. Служат опорой нейронам,обеспеч репаративные процессы,изолир нервн волокно,обеспеч транспорт в-в из капилляров в нейр и наоборот. Олигодендроциты – к-ки,имеющие малое кол-во отростков.меньше по размеру,чем астроциты,участвуют в миелинизации волокон, сод-ся в белом в-ве,в метаболизме нейронов,трофике нейронов.Микроглия-самые мелкие к-ки,многоотросчатые,способны к фагоцитозу,способны изменять размер. Изменение размера глиальных клеток носит ритмический характер и это очень медленный процесс. Частота «пульсации» варьирует от 2 до 20 в час. «Пульсация» происходит в виде ритмического уменьшения объема клетки. Отростки клетки набухают, но не укорачиваются. «Пульсация» усиливается при электрической стимуляции глии; латентный период в этом случае весьма большой — около 4 мин. Глиальные к-ки не обладают импульсной активностью,подобно нервным,однако,мембрана глиальн кл-к имеет заряд,формирующий мембр потенциал,кот отлич большой инертностью. Глиальные к-ки способны передавать возб-е,распространение которого от кл-ки к кл-ке будет ослабевать. Вслед того,что глия тесно контактирует с нейронами,процессы возб-я нервн Эл-тов сказываются на электрич явлениях глиальных эл-тов .это влияние может быть обусловлено тем, что мембр потенциал нейроглии зависит от конц-и ионов K в окр среде: во время возб-я нейрона много K скапливается в нейроне,что приводит к деполяризации клет мембран.
Типы нейронов
А сейчас разберем, какие бывают типы по количеству отростков, и в чем их особенности:
- Униполярные
имеют только один аксон. Это — чувствительные клетки, в которых возникает возбуждение, и они проводят его далее по аксону. - Псевдоуниполярные
. От тела отходит один небольшой “хвостик”, но он ветвится на два — один из них представляет собой аксон, а другой — дендрит. Такие нейроны находятся, в частности, в нервных узлах спинного мозга. - Биполярные
. Как следует из названия, отростка у них два — аксон и дендрит. Их можно найти в органах чувств, например, на сетчатке глаза. - Мультиполярные
. Имеют один аксон и множество дендритов, расходящихся вокруг подобно щупальцам. Как правило, именно так изображают эти клетки в учебниках, и таких клеток большинство.
Важно! Мультиполярные — преобладают в центральной нервной системе (ЦНС). К ним поступают сигналы от множества соседних нейронов — каждый мультиполярный нейрон может быть связан с 1000 других!
Рис. 2. Схема строения нейрона
Функции нейронов
Какие функции есть у нервной ткани? Нервная система, наряду с эндокринной, осуществляет координацию деятельности всего организма. Каждый нейрон является частью цепи в координации того или иного физиологического (или же психического) процесса. Говоря вообще, основная функция нейрона заключается в получении и передаче информации.
Это справедливо в отношении любой клетки рассматриваемой системы, ведь именно этим она и занимается — получает от одних клеток и передает другим информацию в форме нервных импульсов. Однако для различных нейронов выделяют и более специфические функции. Виды нейронов по функциям:
- Афферентные (чувствительные): получают информацию непосредственно от рецепторов, осуществляя взаимодействие между внешним миром и нашей нервной системой.
- Эфферентные (двигательные): отвечают за осуществление конкретных действий — сокращение мышцы, выделение секрета железой.
- Ассоциативные (вставочные): это все “средние” нейроны в цепочке, их может не быть вовсе или быть несколько в одной рефлекторной дуге. Они сосредоточены в ЦНС, отвечают за обработку информации и, говоря грубо, принятие нервной системой решений о действии организма.
Рис. 3. Типы нейронов
Эфферентные (центробежные) нервные волокна
Рефлекс — стереотипная реакция организма в ответ на раздражитель, реализуемая с помощью нервной системы. Вызывающие рефлексы раздражители могут иметь как физическую (механические, электрические, температурные, звуковые, световые и т. п. раздражители), так и химическую природу. Структурной основой рефлекса является рефлекторная дуга, представляющая собой совокупность морфологически взаимосвязанных образований, обеспечивающих восприятие, передачу и переработку сигналов, необходимых для реализации рефлекса.
Безусловный рефлекс.
1>2>3>4>5>6>7
1 Стимул — раздражение рецепторов
2 Рецепторами называют специализированные образования, предназначенные для восприятия клетками или нервной системой различных по своей природе стимулов или раздражителей. Различают два типа рецепторов — сенсорные, т. е. обеспечивающие восприятие нервной системой различных раздражителей внешней или внутренней среды, и клеточные химические рецепторы — специальные структуры мембран, обеспечивающие восприятие информации, переносимой молекулами химических веществ — медиаторов, гормонов, антигенов и т.п.
3 Афферентные (центростремительные) нервные волокна — нервные волокна (отростки нервных клеток ), которые проводят чувствительные импульсы от всех тканей и органов тела к ЦНС . Афферентные нервные волокна (лат. afferens, приносящий) — центростремительные нервные волокна — нервные волокна (отростки нервных клеток), по которым возбуждение передается от тканей к центральной нервной системе. При действии стимулов (влияний среды на органы чувств) в рецепторах возникают потенциалы, которые вызывают возбуждение афферентных сенсорных нервных волокон которое дальше передается в центральную нервную систему . В желудочках сердца расположены рецепторы растяжения . Афферентные волокна от них идут в составе блуждающих нервов . Афферентные волокна от рецепторов растяжения легких также идут в составе блуждающих нервов . Нервные структуры, ответственные за регуляцию водно-солевого баланса , локализованы в промежуточном мозге , особенно в гипоталамусе и соседних областях. Во фронтальной части гипоталамуса расположены многочисленные осморецепторы , которые активируются повышением внутриклеточной концентрации солей при утрате клетками воды и служат чувствительным аппаратом жажды. Кроме того, предполагают, что рецепторы растяжения в стенках крупных вен вблизи сердца также участвуют в регуляции водного баланса и возникновения ощущения жажды при потере воды из внеклеточного пространства. Гипоталамус является важным центром передачи информации от афферентных волокон блуждающего нерва , связанных с рецепторами растяжения, в центральную нервную систему .
4 Нервный центр — это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной функции.
Основными клеточными элементами нервного центра являются многочисленные нейроны, скопление которых формирует нервные ядра. В состав центра могут входить нейроны, рассеянные за пределами ядер. Нервный центр может быть представлен структурами мозга, располагающимися на нескольких уровнях центральной нервной системы (например, центры регуляции дыхания, кровообращения, пищеварения).
Любой нервный центр состоит из ядра и периферии.
Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Повреждение этого участка нервного центра приводит к повреждению или существенному нарушению осуществления данной функции.
Периферическая часть нервного центра получает небольшую порцию афферентной информации, и ее повреждение вызывает ограничение или уменьшение объема выполняемой функции
(рис. 1).
Рис. 1. Схема общего строения нервного центра
Функционирование центральной нервной системы осуществляется благодаря деятельности значительного числа нервных центров, представляющих собой ансамбли нервных клеток, объединенных с помощью синаптических контактов и отличающихся огромным разнообразием и сложностью внутренних и внешних связей.
В нервных центрах выделяют следующие иерархические отделы: рабочие, регуляторные и исполнительные (рис. 2).
Рис. 2. Схема иерархического подчинения разных отделов нервных центров
Рабочий отдел нервного центра ответствен за осуществление данной функции. Например, рабочий отдел дыхательного центра представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту; нарушение этого отдела вызывает остановку дыхания.
Регуляторный отдел нервного центра — это центр, расположенный в коре больших полушарий мозга и регулирующий активность рабочего отдела нервного центра. В свою очередь, активность регуляторного отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию, и от внешних стимулов среды. Так, регуляторный отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной им пульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации углекислого газа и кислорода в крови).
Исполнительный отдел нервного центра — это двигательный центр, расположенный в спинном мозге и передающий информацию от рабочего отдела нервного центра к рабочим органам. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам.
С другой стороны, одни и те же нейроны головного и спинного мозга могут участвовать в регуляции разных функций. Например, клетки центра глотания участвуют в регуляции не только акта глотания, но и акта рвоты. Этот центр обеспечивает все последовательные стадии акта глотания: движение мышц языка, сокращение мышц мягкого неба и его поднятие, последующее сокращение мышц глотки и пищевода при прохождении пищевого комка. Эти же нервные клетки обеспечивают сокращение мышц мягкого нёба и его поднятие во время акта рвоты. Следовательно, одни и те же нервные клетки входят и в центр глотания, и в центр рвоты.
Свойства нервных центров
Свойства нервных центров зависят от их строения и механизмов передачи возбуждения в синапсах. Выделяются следующие свойства нервных центров:
- Односторонность проведения возбуждения
- Синаптическая задержка
- Суммация возбуждения
- Трансформация ритма
- Утомляемость
- Конвергенция
- Дивергенция
- Иррадиация возбуждения
- Концентрация возбуждения
- Тонус
- Пластичность
- Облегчение
- Окклюзия
- Реверберация
- Пролонгирование
Одностороннее проведение возбуждение в нервном центре. Возбуждение в ЦНС проводится в одном направлении с аксона на дендрит или тело клетки следующего нейрона. Основу этого свойства составляют особенности морфологической связи между нейронами.
Одностороннее проведение возбуждения зависит от строения синапса и гуморальной природы передачи в нем импульса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие медиатор, расположены на постсинаптической мембране;
Замедление проведения возбуждения (центральная задержка). В системе рефлекторной дуги медленнее всего проводится возбуждение в синапсах ЦНС. В связи с этим центральное время рефлекса зависит от количества вставочных нейронов.
Чем сложнее рефлекторная реакция, тем больше центральное время рефлекса. Его величина связана со сравнительно медленным проведением возбуждения через последовательно включенные синапсы. Замедление проведения возбуждения создается вследствие относительной длительности осуществляющихся в синапсах процессов: выделения медиатора через пресинаптическую мембрану, его диффузии через синаптическую щель, возбуждения постсинаптической мембраны, возникновения возбуждающего постсинаптического потенциала и его перехода в потенциал действия;
Трансформация ритма возбуждения. Нервные центры способны изменять ритм поступающих к ним импульсов. Они могут отвечать на одиночные раздражители серией импульсов или на раздражители небольшой частоты — возникновением более частых потенциалов действия. В результате ЦНС посылает к рабочему органу количество импульсов, относительно независимое от частоты раздражений.
Это связано с тем, что нейрон является изолированной единицей нервной системы, к нему в каждый момент приходит множество раздражений. Под их влиянием происходит изменение мембранного потенциала клетки. Если создается небольшая, но продолжительная деполяризация (длительный возбуждающий постсинаптический потенциал), то один стимул вызывает серию импульсов (рис. 3);
Рис. 3. Схема трансформации ритма возбуждения
Последействие — способность сохранять возбуждение после окончания действия раздражителя, т.е. афферентных импульсов нет, а эфферентные продолжают действовать еще некоторое время.
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд могут возникать потенциалы действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия.
Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям к первоначально возбужденному нейрону (рис. 4);
Рис. 4. Схема кольцевых связей в нервном центре (по Лоренто де Но): 1 — афферентный путь; 2-промежуточные нейроны; 3 — эфферентный нейрон; 4 — эфферентный путь; 5 — возвратная ветвь аксона
Облегчение проведения или проторение пути. Установлено, что после возбуждения, возникшего в ответ на ритмическое раздражение, следующий стимул вызывает больший эффект, или для поддержания прежнего уровня ответной реакции требуется меньшая сила последующего раздражения. Это явление получило название «облегчение».
Его можно объяснить тем, что при первых стимулах ритмического раздражителя происходит перемещение пузырьков медиатора ближе к пресинаптической мембране и при последующем раздражении медиатор быстрее выделяется в синаптическую щель. Это, в свою очередь, приводит к тому, что вследствие суммации возбуждающего постсинаптического потенциала быстрее достигается критический уровень деполяризации и возникает распространяющийся потенциал действия (рис. 5);
Рис. 5. Схема облегчения проведения
Суммация , впервые описанная И.М. Сеченовым (1863) и заключающаяся в том, что слабые по силе раздражители, не вызывающие видимой реакции, при частом повторении могут суммироваться, создавать надпороговую силу и вызывать эффект возбуждения. Различают два вида суммации — последовательную и пространственную.
- Последовательная суммация в синапсах возникает в том случае, когда по одному и тому же афферентному пути к центрам поступает несколько подпороговых импульсов. В результате суммации местного возбуждения, вызванного каждым подпороговым стимулом, возникает ответная реакция.
- Пространственная суммация заключается в появлении рефлекторной реакции в ответ на два или несколько подпороговых стимулов, приходящих в нервный центр по разным афферентным путям (рис. 6);
Рис. 6. Свойство нервного центра — суммация пространственная (Б) и последовательная (А)
Пространственную суммацию, как и последовательную, можно объяснить тем, что при подпороговом раздражении, пришедшем по одному афферентному пути, выделяется недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны до критического уровня. Если же импульсы приходят одновременно несколькими афферентными путями к одному и тому же нейрону, в синапсах выделяется достаточное количество медиатора, необходимое для пороговой деполяризации и возникновения потенциала действия;
Иррадиация. При возбуждении нервного центра нервные импульсы распространяются на соседние центры и приводят их в деятельное состояние. Это явление получило название иррадиации. Степень иррадиации зависит от количества вставочных нейронов, степени их миелинизации, силы раздражителя. Со временем в результате афферентной стимуляции только одного нервного центра зона иррадиации уменьшается, происходит переход к процессу концентрации, т.е. ограничению возбуждения только в одном нервном центре. Это является следствием уменьшения синтеза медиаторов во вставочных нейронах, в результате чего биотоки не передаются из данного нервного центра на соседние (рис. 7 и 8).
Рис. 7. Процесс иррадиации возбуждения в нервных центрах: 1, 2, 3 — нервные центры
Рис. 8. Процесс концентрации возбуждения в нервном центре
Выражением данного процесса является точная координированная двигательная реакция в ответ на раздражение рецептивного поля. Формирование любых навыков (трудовых, спортивных и т.д.) обусловлено тренировкой двигательных центров, основу которых составляет переход от процесса иррадиации к концентрации;
Индукция. Основой взаимосвязи между нервными центрами является процесс индукции — наведение (индуцирование) противоположного процесса. Сильный процесс возбуждения в нервном центре вызывает (наводит) торможение в соседних нервных центрах (пространственная отрицательная индукция), а сильный тормозной процесс индуцирует в соседних нервных центрах возбуждение (пространственная положительная индукция). При смене этих процессов в пределах одного центра говорят о последовательной отрицательной или положительной индукции. Индукция ограничивает распространение (иррадиацию) нервных процессов и обеспечивает концентрацию. Способность к индукции в значительной степени зависит от функционирования тормозных вставочных нейронов — клеток Реншоу.
От степени развития индукции зависят подвижность нервных процессов, возможность выполнения движений скоростного характера, требующих быстрой смены возбуждения и торможения.
Индукция является основой доминанты — образования нервного центра повышенной возбудимости. Это явление впервые было описано А.А. Ухтомским. Доминантный нервный центр подчиняет себе более слабые нервные центры, притягивает их энергию и за счет этого еще более усиливается. В результате этого раздражение различных рецепторных полей начинает вызывать рефлекторный ответ, характерный для деятельности этого доминантного центра. Доминантный очаг в ЦНС может возникать под влиянием разных факторов, в частности сильной афферентной стимуляции, гормональных воздействий, мотиваций и т.д. (рис. 9);
Рис. 9. Формирование доминанты за счет пространственной отрицательной индукции.
Дивергенция и конвергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками в пределах одного или разных нервных центров называется дивергенциеи. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих вставочных нейронах. Благодаря этому одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов, что приводит к иррадиации возбуждения.
Схождение различных путей проведения нервных импульсов к одному и тому же нейрону получило название конвергенции. Простейшим примером конвергенции является замыкание на одном двигательном нейроне импульсов от нескольких афферентных (чувствительных) нейронов. В ЦНС большинство нейронов получают информацию от разных источников благодаря конвергенции. Это обеспечивает пространственную суммацию импульсов и усиление конечного эффекта (рис. 10).
Рис. 10. Дивергенция и конвергенция
Явление конвергенции было описано Ч. Шеррингтоном и получило название воронки Шеррингтона, или эффекта общего конечного пути. Данный принцип показывает, как при активации различных нервных структур формируется конечная реакция, что имеет первостепенное значение для анализа рефлекторной деятельности;
Окклюзия и облегчение. В зависимости от взаимного расположения ядерных и периферических зон разных нервных центров может проявиться при взаимодействии рефлексов явление окклюзии (закупорки) или облегчения (суммации) (рис. 11).
Рис. 11. Окклюзия и облегчение
Если происходит взаимное перекрывание ядер двух нервных центров, то при раздражении афферентного поля первого нервного центра условно возникают два двигательных ответа. При активации только второго центра также возни каст два двигательных ответа. Однако при одновременной стимуляции обоих центров суммарный двигательный ответ равен только трем единицам, а не четырем. Это обусловлено тем, что один и тот же мотонейрон относится одновременно к обоим нервным центрам.
Если происходит перекрывание периферических отделов разных нервных центров, то при раздражении одного центра возникает одна ответная реакция, то же наблюдается и при раздражении второго центра. При одновременном возбуждении двух нервных центров возникает три ответных реакции. Потому что мотонейроны, находящиеся в зоне перекрывания и не дающие ответа при изолированном раздражении нервных центров, получают при одновременной стимуляции обоих центров суммарную дозу медиатора, что приводит к пороговому уровню деполяризации;
Утомляемость нервного центра. Нервный центр обладает малой лабильностью. Он постоянно получает от множества высоколабильных нервных волокон большое количество стимулов, превышающих его лабильность. Поэтому нервный центр работает с максимальной загрузкой и легко утомляется.
Исходя из синаптических механизмов передачи возбуждения утомление в нервных центрах может объясняться тем, что но мере работы нейрона истощаются запасы медиатора и становится невозможной передача импульсов в синапсах. Кроме того, в процессе деятельности нейрона наступает постепенное снижение чувствительности его рецепторов к медиатору, что называется десенситизацией;
Чувствительность нервных центров к кислороду и некоторым фармакологическим веществам. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходимы энергия и постоянный приток нужного количества кислорода.
Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга, после пяти-шести минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола, их функция восстанавливается через 15-20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливаются даже после 30 мин отсутствия кровообращения.
По сравнению с нервным центром нервное волокно малочувствительно к недостатку кислорода. Помешенное в атмосферу азота, оно только через 1,5 ч прекращает проведение возбуждения.
Нервные центры обладают специфической реакцией на различные фармакологические вещества, что свидетельствует об их специфичности и своеобразии протекающих в них процессов. Например, никотин, мускарин блокируют проведение импульсов в возбуждающих синапсах; их действие приводит к падению возбудимости, уменьшению двигательной активности и полному ее прекращению. Стрихнин, столбнячный токсин выключают тормозящие синапсы, что приводит к повышению возбудимости ЦНС и увеличению двигательной активности вплоть до общих судорог. Некоторые вещества блокируют проведение возбуждения в нервных окончаниях: кураре — в концевой пластинке; атропин — в окончаниях парасимпатической нервной системы. Есть вещества, действующие на определенные центры: апоморфин — на рвотный; лобелии — на дыхательный; кардиазол — на двигательную зону коры; мескалин — на зрительные центры коры и др.;
Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и приспособляемость нервных центров. Это особенно ярко проявляется при удалении разных отделов мозга. Нарушенная функция может восстанавливаться, если были частично удалены какие-то отделы мозжечка или коры больших полушарий. О возможности полной перестройки центров свидетельствуют опыты по сшиванию функционально различных нервов. Если перерезать двигательный нерв, иннервирующий мышцы конечностей, и его периферический конец сшить с центральным концом перерезанного блуждающего нерва, регулирующего внутренние органы, то через некоторое время периферические волокна двигательного нерва перерождаются (вследствие их отделения от тела клетки), а волокна блуждающего нерва прорастают к мышце. Последние образуют в мышце синапсы, свойственные соматическому нерву, что приводит к постепенному восстановлению двигательной функции. В первое время после восстановления иннервации конечности раздражение кожи вызывает свойственную блуждающему нерву реакцию — рвоту, гак как возбуждение от кожи по блуждающему нерву поступает в соответствующие центры продолговатого мозга. Через некоторое время раздражение кожи начинает вызывать обычную двигательную реакцию, поскольку происходит полная перестройка деятельности центра.
Эфферентное волокно
Эфферентные (центробежные) нервные волокна
Эфферентные (центробежные) нервные волокна — нервные волокна, по которым возбуждение передается от ЦНС (от клетки) к тканям. Эфферентные (центробежные) нервные волокна являются отростками эфферентных (двигательных) нейронов .
ЭФФЕКТОР
ЭФФЕКТОР (в физиологии) — конечный элемент рефлекторной дуги (мышца, железа), изменение состояния которого служит показателем осуществления рефлекса (напр., сокращение мышцы, выделение секрета железой). Возможно и нерефлекторное возбуждение эффектора (химическими веществами через кровь, лимфу).
в физиологии эффекторным органом часто называют исполнительный орган или орган-мишень воздействия, выполняющий те или иные «приказы» ЦНС или эндокринных желёз. Например, в случае рефлекторного отдёргивания руки от горячей плиты эффекторным органом является рука. В случае выброса в кровь АКТГ эффекторным органом является кора надпочечников. А в случае вызванного стрессом увеличения концентрации адреналина в плазме крови и увеличения потока импульсов симпатической стимуляции из ЦНС эффекторными органами являются все органы, имеющие симпатическую иннервацию или обладающие адренорецепторами (сердце, бронхи, мышцы и др.). Эффекторным концом (или эффекторным терминалом, эффекторным синапсом) называют дистальный конец аксона, с помощью которого нейрон непосредственно контактирует с органом или тканью, которые он стимулирует или ингибирует.
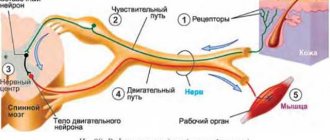
1 . Деятельность организма – закономерная рефлекторная реакция на стимул. Рефлекс – реакция организма на раздражение рецепторов, которая осуществляется с участием ЦНС. Структурной основой рефлекса является рефлекторная дуга (последовательно соединенная цепочка нервных клеток, которая обеспечивает осуществление реакции, ответа на раздражение).
Рефлекторная дуга состоит из шести компонентов: рецепторов, афферентного (чувствительного) пути, рефлекторного центра, эфферентного (двигательного, секреторного) пути, эффектора (рабочего органа), обратной связи.
Рефлекторные дуги могут быть двух видов: простые – моносинаптические рефлекторные дуги; сложные – полисинаптические рефлекторные дуги.
Петля обратной связи – компонент, который устанавливает связь между реализованным результатом рефлекторной реакции и нервным центром, который выдает исполнительные команды. При помощи этого компонента происходит трансформация открытой рефлекторной дуги в закрытую.
Рефлекторная дуга по своему строению и назначению элементов представляет собой описанный выше контур регуляции. Она включает следующие звенья :
1) сенсорные рецепторы (датчики), воспринимающие стимулы внешней или внутренней среды,
2) афферентные, или чувствительные, нервные проводники (каналы сигналов входа),
3) нервные центры (аппарат управления), состоящие из афферентных, промежуточных, или вставочных, и эфферентных нейронов, т. е. получающих, обрабатывающих и выдающих информацию нервных клеток,
4) эфферентные, или двигательные, нервные проводники (каналы выхода),
5) эффекторы, или исполнительные органы (объекты управления).
Однако для оптимальности регуляции необходима информация о реакциях эффектора на управляющие сигналы, в связи с чем обязательным звеном рефлекторного акта является канал обратной связи. Таким образом, структурную основу рефлекса лучше называть не рефлекторной дугой, а рефлекторным кольцом.
Передача нервного импульса
Каким образом импульс передается между клетками? Этот вопрос интереснее, чем кажется на первый взгляд. Сейчас мы поймем, зачем же нейронам сильно развитые ЭПС и аппарат Гольджи. Место контакта аксона с дендритом, телом другого нейрона или эффектором называется синапс. Помимо связи между клетками, синапс также осуществляет перекодировку сигнала, меняя различные его характеристики (частоту, амплитуду). Рассмотрим подробнее схему работы синаптической передачи нервного импульса:
- Электрический нервный импульс возбуждает мембрану нейрона
. Наиболее возбудимая часть на теле таких клеток — аксонный холмик, с него и начинается общее возбуждение клетки. Оно передается далее по мембране аксона и достигает его конца — пресинаптической мембраны. - Внутри аксона содержатся пузырьки с нейромедиаторами
— биологически активными органическими веществами. Помните про ЭПС и аппарат Гольджи? Именно эти системы отвечают за синтез и транспорт веществ-медиаторов к аксонному окончанию. - Синаптическая щель — пространство между аксоном этой клетки и дендритом следующей.
Под действием электрического возбуждения пресинаптическая мембрана высвобождает медиаторы в пространство синаптической щели. Там они связываются с соответствующими рецепторами на постсинаптической мембране, запуская тем самым возбуждение следующего звена нервной цепочки.
Важно! На уровне нейро-физиологии эти процессы выглядят достаточно сложно, и для глубокого понимания требуются знания не только биологии, но и химии, и физики. Однако основное, что нужно понимать — есть две мембраны, связь между которыми осуществляют медиаторы (буквальный перевод — посредники). От того, какой медиатор выделяется, зависит эффект — возбуждение или торможение следующей клетки.
Подведем итоги: нейроны — клетки нервной системы. Их слаженная работа обеспечивает координацию всех функций организма, начиная от движения и работы внутренних органов и заканчивая высшими психическими процессами. Нейроны передают информацию посредством электрических импульсов, которые проходят по аксону одной клетки и при помощи нейромедиаторов в синапсе передаются другой клетке. Такой с виду простой механизм лежит в основе работы нервной системы. Лучше понять работу всей нервной системы вам также поможет предложенное ниже видео.
Предыдущая
АнатомияЖизненная емкость легких, дыхательный объем легких взрослого человека, норма в литрах, таблица, глубина дыхания, виды легочных объемов, функциональная остаточная емкость легких
Следующая
АнатомияКруги кровообращения кратко и понятно, схема артерий, кровеносных сосудов большого и малого круга, последовательность движения крови по венам