Процессы возбуждения в нейропсихиатрии
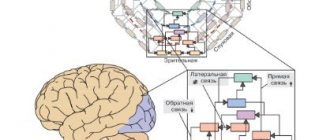
Биомаркеры необходимы для диагностики, выбора и прогноза терапии психоневрологических расстройств. Перспективными направлениями поиска биомаркеров являются дисбаланс возбуждения и торможения в мозге, а также нарушения его созревания.Выявлено два паттерна экспрессии генов: снижение маркеров зрелости и увеличение маркеров незрелости. Эти две группы генов характеризовались избыточной представленностью генов, связанных с синаптической функцией и хромосомной модификацией соответственно. Транскриптомные псевдоиммунные нарушения , индуцируемые нервным гипервозбуждением, разделяется множеством нервно-психических расстройств, например, такими как шизофрения, болезнь Альцгеймера и биполярное аффективное расстройство. Данный эндофенотип служит основой для трансдиагностической характеристики этих нейропсихиатрических расстройств.
Сегодня есть доказательства того, что пациенты с различными клиническими диагнозами имеют сходные биологические особенности, такие как генетические мутации, молекулярная экспрессия или активность мозга. В последнее время в психиатрии происходит «тектонический сдвиг№ в результате ее включения в концепции современной биологии. В последнее время предпринимались неоднократные попытки реклассифицировать психические расстройства в соответствии с биологическими доменами (например, генами, нейронными цепями, поведением), например, в рамках инициативы Research Domain Criteria (RDoC). Следовательно, идентификация биомаркеров, которые могут быть использованы для трансдиагностической оценки нейропсихиатрических расстройств, имеет важное значение для лучшей классификации этих заболеваний и понимания их биологической основы.
Перекрестная экспрессия генов с разными расстройствами может использоваться для характеристики пяти основных психоневрологических расстройств. Некоторые из этих перекрывающихся групп генов были биологически хорошо охарактеризованы обогащением генной онтологии или специфичностью типа клеток, но биологические свойства других групп генов остаются довольно неясными. Таким образом, анализ несмещенной коэкспрессионной сети не обязательно обнаруживает модули, которые выделяют биологические особенности нейропсихиатрических расстройств.
Псевдоиммунитет мозга может быть индуцирован в зрелом возрасте. Ранее было обнаруженно, что длительное лечение флуоксетином полностью изменяет статус созревания нейронов зубчатой извилины (DG ) у взрослых мышей дикого типа — феномен, который можно назвать «дематурацией». Аналогичным образом, недавние исследования предполагают, что некоторые связанные со зрелостью гены и электрофизиологические свойства в DG взрослых мышей дикого типа приобретают незрелый статус после лечения пилокарпином или электросудорожной стимуляцией. IDG-подобный фенотип был обнаружен у пациентов с эпилепсией. Можно предположить, что гипервозбуждение нейронов может быть причиной псевдо-незрелости мозга во взрослом возрасте.
Некоторые исследования предполагают, что гипервозбуждение нейронов может лежать в основе аномалий, связанных с определенными типами психоневрологических расстройств. Пациенты с эпилепсией имеют повышенный риск развития шизофрении, и наоборот и пациенты с эпилепсией также могут демонстрировать психотические симптомы, которые похожи на те, которые обнаруживаются у больных шизофренией. Было высказано предположение, что дисбаланс в возбуждающей и тормозящей схемах головного мозга участвует в патогенезе шизофрении. Гиперактивное возбуждение потенциала действия также наблюдалось в нейронах, подобных гранулярным клеткам гиппокампа, полученных из индуцированных плюрипотентных стволовых клеток пациентов с биполярным расстройством. Последние исследования показали, что пациенты с болезнью Альцгеймера и височной эпилепсией могут иметь общие механизмы , лежащие в основе этих нейропсихиатрических расстройств. Паттерны экспрессии незрелых генов, индуцированные гипервозбуждением нейронов , могут пересекаться с патологическими паттернами экспрессии генов в мозге пациентов с психоневрологическими расстройствами.
Многие генетические риски и факторы окружающей среды, такие как судороги, гипоксия и инфекция, способствуют развитию психоневрологических расстройств. Многие из наборов данных от пациентов с шизофренией и из посмертного мозга пациентов с болезнью Альцгеймера демонстрировали изменения структуры доминантного индекса hiM. Гены hiM включают рецептор ГАМК, потенциал-зависимый кальциевый канал, глутаматный рецептор и потенциал-зависимый натриевый канал. Сообщалось, что эти гены вовлечены в патологические изменения в мозге пациентов с шизофренией и болезнью Альцгеймера. Таким образом, многие из синаптических генов, которые изменились в головном мозге пациентов с шизофренией или болезнью Альцгеймера, могут быть генами, экспрессия которых увеличивается во время созревания и уменьшается при гипервозбуждении нервной системы. Хотя снижение экспрессии некоторых синаптических генов при этих расстройствах хорошо документировано, результаты ряда исследований указывают на возможность того, что гипервозбуждение нейронов может также вызывать сокращения в таких синаптических молекулах.
Большинство наборов данных от пациентов с биполярным аффективным расстройством и болезнью Альцгеймера демонстрировали паттерны доминантного индекса hiI. Гены hiI включают ДНК-метилтрансферазу, циклин, циклин-зависимую киназу, интегрин-бета-3-связывающий белок и опухолевый белок р53. Эти гены , как известно, играют важную роль в хромосомной модификации и репарации ДНК, и аномальные функции этих систем наблюдались у пациентов с биполярным расстройством и при болезни Альцгеймера. Таким образом, некоторые из генов, которые считаются важными в развитии этих расстройств, являются генами, связанными с незрелостью, экспрессия которых уменьшается во время созревания и может быть увеличена за счет гипервозбуждения нейронов.
Что касается наборов данных от пациентов с болезнью Паркинсона (PD), биполярным расстройством (BPD), болезнью Хантингтона (HD) и большим депрессивным расстройством (MDD), у большинства из них не было значительного перекрытия ни генов hiM, ни hiI, что указывает на то, что не может быть патологических изменений псевдоиммунитета, индуцируемых гипервозбуждением нервной системы в наборах данных этих четырех заболевания. Таким образом, можно предположить, что анализ экспрессии генов, основанный на результатах общих эндофенотипов, полезен для проведения трансдиагностической характеристики нервно-психических расстройств.
Возбуждение
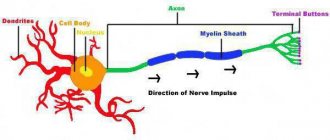
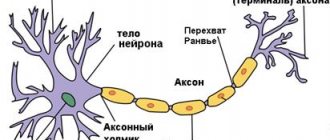
(биологическое), реакция живой клетки на раздражение, выработанная в процессе эволюции. При В. живая система переходит из состояния относительного физиологического покоя к деятельности (например, сокращение мышечного волокна, выделение секрета железистыми клетками и др.). В основе В. лежат сложные физико-химические процессы. Начальный пусковой момент В. — изменения ионной проницаемости и электрических потенциалов мембраны. Наиболее полно В. изучено в нервных и мышечных клетках, где оно сопровождается возникновением потенциала действия (ПД), способного без затухания (бездекрементно) распространяться вдоль всей клеточной мембраны (см.
Биологические мембраны
). Это свойство ПД обеспечивает быструю передачу информации по периферическим нервам в нервные центры и от них к исполнительным органам — мышцам и железам. В волокнах скелетных мышц ПД распространяется как вдоль мембраны, так и в глубь волокна к сократительному аппарату миофибрилл; поэтому вслед за волной В. по мышечному волокну распространяется волна сокращения. Пусковое влияние ПД оказывает и на секрецию нервными окончаниями химических веществ —
медиаторов
, оказывающих возбуждающее или тормозящее действие на иннервируемые ткани. ПД подчиняется правилу
«всё или ничего»
: он возникает только после достижения раздражителем пороговой величины (порог раздражения) и сразу приобретает максимальную амплитуду. Во время развития ПД клетка полностью утрачивает
возбудимость
, т. е. способность отвечать новым возбуждением на повторный стимул. Возбудимость восстанавливается постепенно лишь после окончания ПД (см.
Рефрактерный период
).
Распространяющийся импульс — не единственная форма В. В определённых участках нервных и мышечных клеток позвоночных животных и в некоторых клетках беспозвоночных В. имеет местный характер. Среди разновидностей местного В. наиболее важное функциональное значение имеют генераторные потенциалы рецепторов и возбуждающие постсинаптические потенциалы, возникающие в области контакта клетки с двигательными нервными окончаниями (см. Синапсы
). Так же как и ПД, местное В. связано с избирательным повышением ионной проницаемости мембраны и проявляется в виде отрицательного колебания поверхностного потенциала (см.
Деполяризация
). Однако, в отличие от ПД, местное В. не подчиняется правилу «всё или ничего»: оно не имеет порога и варьирует по амплитуде и длительности в зависимости от силы и длительности раздражителя. Деполяризация мембраны, сопровождающая местное В., служит раздражителем для соседних участков мембраны, способных к генерации распространяющегося ПД. Поэтому при достижении местным В. (генераторным или постсинаптическим потенциалом) пороговой величины возникает ПД. Различия в свойствах местного и распространяющегося В. имеют важное значение для процессов передачи информации нервными клетками и их волокнами. Местное В. свойственно тем участкам клеточной мембраны, которые специализированы на восприятии раздражений, приходящих извне (рецепторная мембрана) или от других нервных клеток (постсинаптическая мембрана). Местное В. имеет градуальный характер и потому может более тонко отражать характеристики раздражителя — его силу, длительность, скорость нарастания и падения, — чем ПД, возникновение которого служит лишь сигналом достижения раздражителем пороговой величины. С другой стороны, способность ПД к быстрому бездекрементному распространению делает его наиболее адекватным для передачи информации по длинным проводникам. При этом информация о силе, длительности и крутизне изменений раздражителя кодируется частотой нервных импульсов, изменением этой частоты во времени и длительностью всего залпа ПД.
Механизм генерации и проведения нервных импульсов, совершенствуясь в процессе эволюции организмов, достиг наивысшего развития у теплокровных животных и человека. Скорость проведения В. в мякотных (миелинизированных) волокнах теплокровных достигла 100—120 м/сек
при длительности ПД 0,2— 0,4
м/сек
. У холоднокровных позвоночных животных (например, земноводных) скорость проведения ПД по мякотным волокнам такого же диаметра (20
мкм
) не превышает 20—30
м/сек
при длительности ПД 1,5—2
м/сек
. В. — комплексная реакция, в которой существенную роль наряду с электрическими играют и структурные, химические (в том числе ферментативные), физические (температурные) и другие компоненты. Изменения мембранного потенциала при В. — следствие избирательного повышения проницаемости мембраны (в результате её молекулярной перестройки) для вне и внутриклеточных ионов (см.
Мембранная теория возбуждения
). Предполагают, что с этим связаны также усиление теплопродукции во время восходящей фазы ПД и поглощение тепла в нисходящую фазу. Проникновение ионов Na+ и (или) Ca2+ в протоплазму во время В. активирует ферментативные процессы, направленные на восстановление исходного неравенства концентраций ионов Na+, К+, Ca2+ по обе стороны мембраны (см.
«Натриевый насос»
) и на синтез белков и фосфолипидов, необходимых для постоянного обновления структуры мембраны и протоплазмы клетки. Активация обменных процессов сопровождается увеличением потребления тканью кислорода и новым повышением теплопродукции, длящимся в нерве в течение многих минут после прохождения импульса. Ср.
Торможение
.
Лит.:
Физиология человека, под ред. Е. Б. Бабского, 2 изд., М., 1936; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Катц Б., Нерв, мышца, синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969.
Б. И. Ходоров.
Оглавление