Классификация нервных волокон
Классификация нервных волокон по диаметру и скорости проведения
− классификация двигательных и сенсорных нервных волокон по Эрлангеру-Гассеру;
− классификация сенсорных нервных волокон по Ллойду-Ханту;
− сравните классификации нервных волокон по Эрлангеру
− Гассеру и Ллойду-Ханту с классификацией нервных волокон по De Meyer (таблицы 1, 2, 3).
Типы нервных волокон и их функции
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 году обнаружили составной характер тока действия смешанного нерва.
На основании полученных данных (диаметр, скорость проведения, функция) была разработана классификация нервных волокон (табл. 1, сводная табл. 3), в соответствии с которой все нервные волокна подразделены на группы А, В и С (латинские буквы) с дальнейшими градациями (α, β и т. д.).
Классификация нервных волокон по Эрлангеру – Гассеру — классификация двигательных и сенсорных нервных волокон:
А, В, С — группы волокон;
α, β, γ, δ — подгруппы волокон группы А.
Волокна типа А – толстые миелиновые нервные волокна.
Наиболее толстые из них Аα имеют диаметр 12 — 22 мкм и скорость проведения возбуждения 70—120 м/с. Эти волокна проводят возбуждение от спинальных моторных нервных центров (двигательных центров спинного мозга) к скелетным мышцам (двигательные волокна) и от рецепторов мышц к соответствующим нервным центрам.
Другие группы волокон типа А (β, γ, δ) имеют меньший диаметр — от 8 до
1 мкм и меньшую скорость проведения возбуждения — от 5 до 70 м/с. Волокна этих групп преимущественно проводят возбуждение от различных рецепторов (тактильных, температурных, болевых; рецепторов внутренних органов или висцерорецепторов) в ЦНС, за исключением γ-волокон, значительная часть которых проводит возбуждение от спинного мозга к интрафузальным мышечным волокнам.
Таблица 1
Классификация нервных волокон по Эрлангеру-Гассеру
| Тип волокон | Функция (выборочно) | Средний диаметр, мкм | Средняя скорость проведения, м/с |
| Аα | Первичные афференты мышечных веретён, двигательные волокна скелетных мышц | 15 | 100 (70 – 120) |
| Аβ | Кожные афференты прикосновения и давления | 8 | 50 (30 – 70) |
| Аγ | Двигательные волокна мышечных веретён | 5 | 20 (15 – 30) |
| Аδ | Кожные афференты температуры и боли | <3 | 15 (12 – 30) |
| В | Симпатические преганглионарные волокна | 3 | 7 (3 – 15) |
| С | Кожные афференты боли Симпатические постганглионарные волокна | 1 (немиелинизированные) | 1 (0,5 – 2) |
К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы. Их диаметр — 1 — 3,5 мкм, а скорость проведения возбуждения — 3—18 м/с.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра — 0,5 — 2 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5 — 3 м/с).
Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Классификация нервных волокон Ллойда-Ханта
Классификация Ллойда-Ханта широко применяется для сенсорных нервных волокон (римские цифры), которая представлена в таблице 2 и сводной таблице 3.
Таблица 2
Классификация нервных волокон по Ллойду-Ханту
| Тип волокон | Функция (выборочно) | Средний диаметр, мкм | Средняя скорость проведения, м/с |
| I | Первичные афференты мышечных веретён и от сухожильных органов | 13 | 75 (70 – 120) |
| II | Кожные механорецепторы | 9 | 55 (25 – 70) |
| III | Мышечные сенсоры глубокого давления | 3 | 11 (10 – 25) |
| IV | Немиелинизированные афференты боли | <3 |
Физиология нервно-мышечной ткани
Известно, что под влиянием раздражителя живые клетки и ткани из состояния физиологического покоя переходят в состояние активности. Наибольшая ответная реакция среди тканей на раздражение наблюдается со стороны нервной и мышечной ткани. Основными свойствами нервной и мышечной ткани являются возбудимость, возбуждение, проводимость, рефрактерность и сократимость.
Возбудимость — это способность живой ткани отвечать на действие раздражителя изменением физиологических свойств и возникновением процесса возбуждения. Возбуждение — это активный физиологический процесс, который возникает в ткани под влиянием раздражителей и характеризуется изменением уровня обменных процессов в тканях, выделением энергии, сокращением мышечной ткани, выделением секрета, генерацией нервного импульса.
Проводимостью называют способность живой ткани проводить волны возбуждения (биопотенциалы). Рефрактерность — это временное снижение возбудимости ткани, которое возникает в результате возбуждения. Лабильность — это зависимость ткани от особенностей обменных процессов, которая может возбуждаться определенное количество раз за единицу времени.
Различают электрические, химические, механические и температурные раз- дражители, которые способны вызвать ответную реакцию со стороны возбудимых тканей. По биологическим признакам раздражители могут быть адекватными и неадекватными, а по силе — подпороговымя, пороговыми и надпороговыми.
Решающее значение для появления возбуждения принадлежит силе раздражителя (закон раздражения). Существует определенная зависимость между силой раздражения и ответной реакцией. Чем больше сила раздражителя, тем выше, до соответствующего уровня, ответная реакция со стороны возбудимой ткани.
Большое значение имеет и продолжительность действия раздражителя. Зависимость между силой раздражителя и продолжительностью его действия, необходимого для появления минимальной ответной реакции, определяется кривой силы — времени. Минимальная сила тока (напряжения), способная вызвать возбуждение, называется реобазой (отрезок ординаты (ОА). Кроме реобазы, важным параметром кривой силы является хроноксия.
Последняя отражает наименьший промежуток времени, во время которого ток, равный по силе удвоенной реобазе (отрезок абсциссы OF), вызывает в ткани возбуждение. По величине хроноксии судят о скорости появления возбуждения в ткани: чем меньше хроноксия, тем быстрее возникает возбуждение.
Приспособление возбудимой ткани к медленному нарастанию силы раздражителя получило название аккомодации. Последняя обусловлена тем, что за время нарастания силы раздражителя в ткани происходят активные изменения, которые повышают порог раздражения и препятствуют развитию возбуждения. Таким образом, скорость нарастания раздражения во времени называют градиентом раздражения.
Закон градиента раздражения — это реакция на раздражитель, которая за- висит от срочности или крутизны нарастания раздражителя за определенное время: чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого объекта.
Потенциал покоя (мембранный потенциал) — это разность потенциалов между наружной поверхностью клетки и ее внутренним содержимым; он составляет около 60—90 мВ в зависимости от особенности той или иной клетки.
Потенциал действия (потенциал возбуждения) возникает при воздействии на участок нервного или мышечного волокна раздражителя достаточной силы и продолжительности. Проявлением возникшего возбуждения является быстрое колебание мембранного потенциала. При этом возбужденный участок имеет электроотрицательный заряд по отношению к невозбужденному.
В потенциале действия различают местные колебания мембранного потенциала, пик потенциала действия и следовые потенциалы — отрицательный и положительный. Пик потенциала действия представляет собой кратковременное изменение внутреннего потенциала и имеет очень быструю восходящую фазу и несколько замедленный спад.
Вслед за пиком потенциала действия регистрируются более слабые и продолжительные отрицательные, а затем положительные следовые потенциалы. Продолжительность потенциала действия в нервных и мышечных волокнах составляет 0,1—5,0 мс. В развитии потенциала покоя и потенциала действия роль играет система проникающих каналов для ионов Na+, К+, С1-, Са2+. В нервной мембране присутствуют специфические натриевые, калиевые, хлорные и кальциевые каналы, которые пропускают только вышеназванные ионы.
Эти каналы обладают воротными механизмами и могут быть открытыми и закрытыми. Определение состояния ионных каналов мембраны очень важно для образования потенциала покоя, где ведущая роль принадлежит неравномерному распределению ионов калия. В образовании потенциала действия основную роль играют ионы натрия. Проведение возбуждения — специализированная функция нервных волокон.
Скорость возбуждения по ним зависит в основном от диаметра и гистологических особенностей строения нервных волокон. Чем больше диаметр нервного волокна, тем выше в нем скорость распространения возбуждения. Например, по нервному волокну диаметром 12— 22 мкм скорость распространения возбуждения составляет 70—120 м/с, а по нервному волокну диаметром 8—12 мкм — только 40—70 м/с.
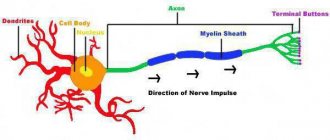
По гистологическому строению нервные волокна делят на миелиновые и безмиелиновые. Миелиновое волокно состоит из осевого цилиндра и покрывающей его миелиновой, или шванновской, оболочкой. В ее состав входят жировидные вещества, которые обладают высоким удельным сопротивлением и выполняют изолирующую роль. Через равные промежутки миелиновая оболочка прерывается и оставляет открытые участки осевого цилиндра шириной около 1 мкм (перехват Ранвье).
Поверхность осевого цилиндра представлена плазматической мембраной, а его содержимое — аксоплазмой. Безмиелиновые волокна не имеют миелиновой оболочки, а покрыты только шванновскими клетками. Пространство между шванновскими клетками и осевым цилиндром заполнено межклеточной жидкостью, что дает возможность поверхностной мембране осевого цилиндра сообщаться с окружающей нервное волокно средой.
Передача возбуждения по миелиновым и безмиелиновым волокнам имеет свои особенности. Так, передача потенциала действия по миелиновому волокну происходит скачкообразно от одного перехвата Ранвье к другому, что дает возможность возбуждению распространяться без угасания.
Скорость распространения по миелиновым волокнам значительно выше, чем по безмиелиновым. Если скорость возбуждения по двигательным нервным волокнам (покрытым миелиновой оболочкой) составляет 80—120 м/с, то по волокнам, которые не имеют миелиновых волокон, — только 0,5—2,0 м/с. Распространение возбуждения по нервному волокну при нанесении возбуждения подчиняется соответствующим законам.
Закон физиологической целостности — проведение возбуждения по нерв- ному волокну возможно только в том случае, если сохраняется не только его ана- томическая, но и физиологическая целостность (непрерывность).
Закон двухстороннего проведения возбуждения — передача возбуждения происходит в двух направлениях — центростремительном и центробежном.
Закон изолированного проведения возбуждения — при нанесении раздражения возбуждение проводится только по одному нервному волокну и не охватывает соседние волокна, что обусловливает строгую координацию рефлекторной деятельности. Нервные волокна мало устают. Это объясняется низкими энергетическими затратами и быстрыми восстановительными процессами.
Синапс — это специализированная структура, которая обеспечивает переда- чу нервного импульса из нервного волокна на эффекторную клетку — мышечное волокно, нейрон или секреторную клетку.
Синапсы классифицируют по анатомо-гистологическому принципу (нейро- секреторные, нервно-мышечные, межнейронные); нейрохимическому принципу (адренергические — медиатор норадреналин и холинергические — медиатор ацетилхолин); функциональному (возбуждающие и тормозные). Нервно-мышечный синапс состоит из трех основных структур: пресинаптической мембраны, синаптической щели и постсинаптической мембраны.
Пресинаптическая мембрана покрывает нервное окончание, а постсинаптическая — эффекторную клетку. Между ними находится синаптическая щель. Постсинаптическая мембрана отличается от пресинаптической тем, что имеет белковые хеморецепторы, чувствительные не только к медиаторам, гормонам, но и к лекарственным и токсическим веществам. Строение нервно-мышечного синапса обусловливает его физиологические свойства:
- односторонее проведение возбуждения (от пре- к постсинаптической мембране) при наличии чувствительных к медиатору рецепторов только в постсинаптической мембране;
- синаптическая задержка проведения возбуждения, связанная с малой скоростью диффузии медиатора в сравнении со скоростью нервного импульса;
- низкая лабильность и высокая усталость синапса;
- высокая избирательная чувствительность синапса к химическим веществам.
Передача возбуждения в синапсе представляет собой сложный физиологический процесс, который проходит несколько стадий:
- синтез медиатора;
- секреция медиатора;
- взаимодействие медиатора с рецепторами постсинаптической мембраны;
- инактивация (полная утрата активности) медиатора.
Известно, что некоторые химические вещества, в том числе и лекарственные, могут существенно влиять на возбуждение в синапсе. Это явление нашло применение в клинической практике.
Нейромоторная единица — это анатомическая и функциональная единица скелетных мышц, которая состоит из аксона (длинного отростка мотонейрона спинного мозга) и иннервируемых им определенного количества мышечных волокон. В состав нейромоторной единицы может входить разное количество мышечных волокон (от единиц до нескольких тысяч), которое зависит от специализации мышцы. Двигательная единица работает как единое целое. Импульсы, выработанные мотонейроном, приводят в действие все образующие ее мышечные волокна.
Основной функцией скелетных мышц является сокращение, которое выражено различными движениями человека. Скелетные мышцы выполняют также рецепторную, обменную и терморегулирующую функции. Они образуются большим количеством многоядерных мышечных волокон.
Сократительной частью мышечного волокна являются длинные мышечные нити — миофибриллы, которые про- ходят внутри волокна от одного конца к другому и имеют поперечную очерченность. Последняя образована чередованием темных (анизотропных) А-дисков и светлых (изотропных) 1-дисков. Через середину 1-диска проходит Z- линия; две соседние Z-линии ограничивают саркомер, структурно-фунциональную единицу. При электронной микроскопии волокон видно, что в составе А-диска есть более светлый участок (Н-зона), а в центре этот диск пересекает темная поло- са — М-линия.
Темный диск образован толстыми нитями белка миозина, а свет- лый 1-диск — тонкими нитями белка актина. В мышечном волокне содержатся также фибриллярный палочковидный белок — тропомиозин и глобулярный белок — тропонин. Механизм сокращения состоит в перемещении (перетягивании) тонких нитей вдоль толстых к центру саркомера за счет поперечных актиномиозино- вых мостиков. Основным источником энергии, необходимой для мышечного со- кращения, служат аденозинтрифосфорная кислота (АТФ) и присутствие ионов Са2+ и Mg2+ .
Преобразование химической энергии в механическую происходит в мышце как без кислорода, так и с его участием. Анаэробная (бескислородная) фаза характеризуется рядом последовательных реакций, которые приводят к распаду АТФ и креатинфосфата, и их восстановлением. Выделенная при этом энергия используется для сокращения мышц и восстановления (ресинтеза) этих веществ. Аэробная (кислородная) фаза химических преобразований связана с процессами окисления молочной кислоты до углекислого газа и воды. Возникшая энергия используется для дальнейшего преобразования остатков молочной кислоты в глюкозу, а затем в гликоген.
Деятельность скелетной мускулатуры регулируется ЦНС — корой головно- го мозга, через чувствительные, двигательные и симпатические нервные волокна. Скелетные мышцы обладают следующими физиологическими свойствами: возбудимостью, проводимостью, рефрактерностью, лабильностью и сокращением. Возбудимость, скорость распространения возбуждения, лабильность мышечной ткани ниже, чем в нервной, а рефракторный период продолжительнее нервного.
Скелетные мышцы могут выполнять работу в режиме изотонического, изометрического и ауксото-ничного сокращения. При первом сокращении в основном происходит укорочение мышечного волокна, но напряжение остается постоянным, а при втором — длина мышечных волокон остается неизменной, но изменяются длина и напряжение. Характер сокращения скелетных мышц зависит от частоты раздражения (частоты поступления нервных импульсов).
Раздражение одиночным импульсом ведет к одиночному мышечному со- кращению, а следующих один за другим нервных импульсов — к тетаническому сокращению, или тетанусу.
Физиологические свойства гладких мышц связаны с особенностью их строения, уровнем обменных процессов и значительно отличаются от особенностей скелетных мышц. Гладкие мышцы менее возбудимы, чем поперечнополосатые. Сокращение гладкой мускулатуры происходит медленнее и продолжительнее. Рефракторный период у гладких мышц более удлиненный, чем у скелетных (до нескольких секунд).
Характерная особенность гладких мышц — их способность к автоматической деятельности, которая обеспечивается нервными элементами. Гладкие мышцы иннервируются симпатическими и парасимпатическими вегетативными нервами, обладают высокой чувствительностью к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и др.).
Миелинизация
Нервное волокно имеет в основе отросток нейрона, который образует своеобразную ось. Снаружи он окружен миелиновой оболочкой с биомолекулярной липидной основой, состоящей из большого количества витков мезаксона, который по спирали накручивается на нейроновую ось. Таким образом, происходит миелинизация нервных волокон.
Миелиновые нервные волокна периферической системы сверху дополнительно покрыты вспомогательными Шванновскими клетками, поддерживающими аксон и питающими тело нейрона. Поверхность мякотной мембраны имеет интервалы – перехваты Ранвье, в этих местах осевой цилиндр прикрепляется к наружной Шванновской мембране.
Миелиновый слой не обладает электропроводящими свойствами, их имеют перехваты. Возбуждение происходит в ближайшем к месту воздействия внешнего раздражителя интервале Ранвье. Импульс передается скачкообразно, от одного перехвата к другому, это обеспечивает высокую скорость распространения импульса.
Миелиновые нервные волокна регулируют обмен веществ в мышечной ткани, обладают высоким сопротивлением по отношению к биоэлектрическому току.
Промежутки Ранвье генерируют и усиливают импульсы. У волокон центральной нервной системы нет Шванновской мембраны, эту функцию выполняют олигодендроглии.
Безмякотные ткани имеют несколько осевых цилиндров, у них нет миелинового слоя и перехватов, сверху покрыты Шванновскими клетками, между ними и цилиндрами образуются щелевидные пространства. Волокна имеют слабую изоляцию, допускают распространение импульса из одного отростка нейрона в другой, на всем протяжении контактируют с окружающей средой, скорость проведения импульсов гораздо ниже, чем у мякотных волокон, при этом организму требуется большее количество энергии.
Из мякотных и безмякотных отростков нейронов формируются крупные нервные стволы, которые, в свою очередь, разветвляются на более мелкие пучки и заканчиваются нервными окончаниями (рецепторные, двигательные, синапсы).
Нервные окончания – это конец миелиновых и безмиелиновых нервных волокон, который формирует межнейронные контакты, рецепторные и двигательные окончания.
Законы проведения возбуждения в нервных волокнах и нервах
Классификация нервных волокон по Эрлангеру-Гассеру
В 1939 г. американские[Мф54] физиологи Джозеф Эрлангер [Б55] и Герберт С.Гассер [Б56] [Б57] зарегистрировали токи действия от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрода (рис. 210041905).[Б58]
Было установлено, что регистрируемый суммарный потенциал имеет ряд пиков, которые были обозначены буквами латинского алфавита A, B, C (рис. , I). Пик A имел дополнительные пики, помеченные греческими буквами α, β, γ, δ (рис. , II). [Б59] В 1944 г. работа Дж.Эрлангера и Г.С.Гассера была оценена присуждением Нобелевской премии[Б60] .
Нервы у позвоночных состоят из трех основных групп волокон
(А, В и С), различающихся по степени миелизации, диаметру волокна, длительности пика ПД (скорости развития ПД), электровозбудимости, его компенсации и скорости проведения (все эти показатели в ряду А — В — С падают).
Группа А
включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна;
группа В —
слабомиелинизированные, преганглионарные волокна автономной нервной системы;
группа С —
немиелинизированные, постганглионарные (симпатические) волокна.
В группе А в ряду a, b, g, d названные показатели тоже падают. Соотношения свойств этих групп волокон демонстрируются в табл. .
Необходимо заметить, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Относительно высокие пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким входным сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для Аa
-волокон она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. По этой же причине (высокое
RI
) отводимые от ствола (внеклеточно) ПД тонких волокон предстают значительно меньшими, чем ПД толстых волокон.
Рис. . Составные части потенциала действия смешанного нерва.
I – при относительно медленной скорости записи.
II – при относительно высокой скорости записи.
Объяснение в тексте. По оси абсцисс – время, по оси ординат амплитуда составного потенциала в мВ.
Суммарная электрическая активность нерва
создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств.
Рассмотрим случай так называемого однофазного отведения,
при котором один отводящий электрод (активный) расположен на нормальном участке ствола, а другой (индифферентный) — на поврежденном, где волокна деполяризованы полностью (рис.1.19). Пусть применяют искусственное раздражение и раздражающий электрод (катод) находится достаточно близко
(l=
3 мм) от активного отводящего электрода, а нерв помещен в непроводящую среду (масло или воздух). Здесь сильное одиночное раздражение приводит к синхронному возбуждению всех волокон, при этом активный отводящий электрод регистрирует суммарный ПД нерва, по форме приближающийся к ПД отдельного A(альфа)‑волокна, но немного более затянутый во времени. Этот ПД нерва, однако, не подчиняется правилу «все или ничего». При пороговом раздражении он ничтожно мал, с увеличением силы стимулов постепенно растет, достигая максимума, равного при обычных условиях 5—10 мВ, а в условиях сахарозного мостика — 50—100 мВ.
При дальнейшем увеличении силы стимула этот ПД несколько удлиняется во времени. Все изменения амплитуды и длительности пика ПД нерва при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым А(альфа)-волокнам более высокопороговых медленных бета-, гамма-, дельта-волокон группы А, затем В- и, наконец, С-группы.
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Скорость проведения, м/с |
| Aa | 13 — 22 | 70 — 120 |
| Ab | 8 -13 | 40 — 70 |
| Ag | 4 — 8 | 15 – 40 |
| Ad | 1 – 4 | 5 – 15 |
| B | 1 — 3 | 3 – 14 |
| C | 0,5 – 1,0 | 0,5 — 2 |
Таблица . Классификация нервных волокон по Дж.Эрлангеру и Х.Гассеру
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Пороги электрического раздраженния (относительно Aa) | Длительность пика ПД1 | Отрицательный следовой потенциал (ОСП) | Положительный следовой потенциал [Б61] | Скорость проведения, м/с | |
| Длительность, мс | Амплитуда СП, % к амплитуде ПД | Длительность, мс | Амплитуда СП, % к амплитуде ПД | ||||
| Aa | 13 — 22 | 1,0 | 0,4 | 15 – 20 | 40 — 60 | 0,2 | 70 — 120 |
| Ab | 8 -13 | 40 — 70 | |||||
| Ag | 4 — 8 | 15 – 40 | |||||
| Ad | 1 – 4 | 5 – 15 | |||||
| B | 1 — 3 | 11,7 | 1,2 | ОСП нет | 100 – 300 | 3 – 14 | |
| C | 0,5 – 1,0 | 100,0 | 2,0 | 50 – 60 | 300 — 1000 | 0,5 — 2 |
1Приблизительно ту же величину имеют и абсолютные рефрактерные фазы
Основные свойства автоволн, касающиеся их распространения, распространяются и на потенциалы действия нервных волокон:
1. распространяется без затухания как по длине волокна, так и при его разветвлении (рис. 709170042).
2. не интерферируют (рис. 709170043).
3. не отражаются от препятствий (рис. 709170044).
4. направление распространения определяется зонами рефрактерности и покоя, обеспечивается двустороннее проведение возбуждения (рис. 709170045, 709170046).
Рис. 709170042. Распространение ПД при разветвлении нервных волокон. Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
Рис. 709170043. Распространение ПД при схождении нервных волокон. Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
Рис. 709170044. ПД не отражаются от препятствий Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
ПД проходит равные расстояния (L) от места действия стимула за одно и то же время (t) и сохраняет амплитуду при прочих равных условиях.
Рис. 709170045. Двустороннее проведение по нервным волокнам. L
– расстояние от места действия стимула,
t
– время проведения ПД от места действия стимула до места расположения регистрирующих электродов,
A
– амплитуда ПД.
Обычно подчёркивается условие сохранения анатомической и физиологической непрерывности волокна
Анатомическая и физиологическая непрерывность волокна
Проведение импульсов возможно лишь при условии анатомической целостности волокна, поэтому как перерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна блокада натриевых каналов возбудимой мембраны тетродотоксином или местными aнестетиками, резкое охлаждение и т.п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна К+, накапливающимися при ишемии в межклеточных щелях. Механическая травма, сдавливание нерва при воспалительном отеке тканей могут сопровождаться частичным или полным нарушением функции проведения.