23.09.2020 Алёна Машева Здоровье
Таламус — это структура головного мозга, которая во внутриутробном развитии формируется из промежуточного мозга, составляя основную его массу у взрослого человека. Именно через это образование вся информация с периферии передается к коре. Второе название таламуса — зрительные бугры. Подробнее о нем далее в статье.
Расположение
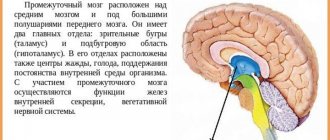
Таламус является частью больших полушарий переднего мозга. Он расположен латеральней боковых желудочков — полостей мозга, которые являются частью системы циркуляции ликвора (спинномозговой жидкости). Снизу от него находится гипоталамус, от которого зрительные бугры отделены бороздой.
Выше и несколько снаружи от таламуса находятся базальные ядра. Эти образования необходимы для осуществления точных, координированных движений. Друг от друга эти структуры разделены внутренней капсулой — пучком белого вещества переднего мозга, через который проходят проводящие пути от периферии к центру.
Между собой правая и левая части таламуса соединены межталамическим серым веществом. Оно присутствует у 70% людей.
Классификация ядер таламуса
Всего в зрительных буграх мозга насчитывается около 120 ядер. В зависимости от места нахождения их подразделяют на три группы:
- медиальные;
- латеральные;
- передние.
- специфические;
- ассоциативные;
- неспецифические.
В латеральной группе ядер выделяют, в свою очередь, медиальное и латеральное коленчатые тела, а также подушку.
Существует также классификация в зависимости от выполняемой функции ядрами:
Специфические ядра
Специфические ядра зрительного бугра имеют ряд отличительных особенностей. Все образования этой группы получают сенсорную информацию от вторых нейронов (нервных клеток) чувствительных проводящих путей. Второй нейрон, в свою очередь, может быть расположен в спинном мозге или в одной из структур ствола мозга: продолговатом мозге, мосту, среднем мозге.
Каждый из сигналов, поступающих снизу, обрабатывается в таламусе и далее идет в соответствующую область коры. В какую именно область поступает нервный импульс, зависит от того, какую информацию он несет. Так, информация о звуках поступает в слуховую кору, об увиденных предметах — в зрительную и так далее.
Помимо импульсов из вторых нейронов проводящих путей, специфические ядра отвечают за восприятие информации, поступающей из коры, ретикулярной формации, ядер ствола мозга.
Ядра, которые находятся в передней части таламуса, обеспечивают проведение импульсов из лимбической коры головного мозга через гиппокамп и гипоталамус. После обработки информации она опять поступает в лимбическую кору. Таким образом, нервный импульс циркулирует по определенному кругу.
Таламус
Определение понятия
Таламус (thalamus opticus — зрительный бугор) — это отдел промежуточного мозга, управляющий потоками сенсорного возбуждения, идущими к нему от всех органов чувств. Его основные функции: трансформация сенсорного возбуждения, афферентное взаимодействие с корой, лимбической системой, стрио-паллидарной системой, гипоталамусом, а также обеспечение внимания.
«Запоминалка»
«Таламический бугор — ощущениям отбор». Таламус — как личный секретарь, который получает всю информацию, но передаёт своему начальнику только самую важную и в сжатом и понятном виде, а затем передаёт распоряжения начальника исполнителям.
Таламус («зрительный бугор») обеспечивает подготовку сенсорного возбуждения, приходящего от органов чувств, для передачи в определённые зоны коры больших полушарий головного мозга. Таламус фильтрует информацию, поступающую от всех рецепторов, осуществляет её предварительную обработку и после этого направляет её в соответствующие области коры. Кроме того, таламус осуществляет связь между корой, с одной стороны, и мозжечком и базальными ганглиями с другой. Иными словами, через таламус низшие нервные центры отчитываются перед высшими, а высшие корковые нервные центры управляют работой низших нервных центров.
Строение таламуса
Таламус относится к промежуточному мозгу, который расположен между средним мозгом и большими полушариями переднего мозга. Он состоит из 40 ядер. Можно сказать, что таламус занимает центр мозга, и это соответствует его центральной роли в обработке информации, получаемой мозгом.
Таламус собирает сенсорное возбуждение, приходящее по афферентным путям от внешних экстерорецепторов и внутренних интеррецепторов и готовит его для передачи в кору, а затем передаёт его в разные зоны коры по разным афферентным путям: специфическим, неспецифическим и ассоциативным. Только обонятельное сенсорное возбуждение приходит в таламус из обонятельной коры, все остальные сенсорные потоки вначале попадают в таламус, а затем из него — в кору.
При повреждении таламуса кора может лишиться той или иной сенсорной информации и сенсорное восприятие нарушится.
Ядра таламуса делятся на специфические и неспецифические. Соответственно, и пути от них к коре больших полушарий делятся на специфические и неспецифические. Специфические ядра, в свою очередь, делятся на переключательные и ассоциативные. Характеристика ядер. Специфические. Делятся на переключательные и ассоциативные. Переключательные. Осуществляют переключение потока сенсорного возбуждения от низших нервных центров спинного мозга и ствола в сенсорные зоны коры. Предварительно происходит перекодирование и обработка полученного сенсорного возбуждения. Вентральное переднее. Регуляция движений. Вентральное заднее.Переключают соматосенсорную афферентную информацию: тактильную, проприоцептивную, вкусовую, висцеральную, частично температурную, болевую. Латеральное коленчатое тело.Переключение зрительной информации в затылочную область коры. Медиальное коленчатое тело.Переключение слуховой информации в височную кору задней части сильвиевой борозды (извилины Гешля). Ассоциативные. Получают афферентные сигналы от переключающих ядер и направляют в ассоциативные зоны коры. Главная функция — интеграция деятельности таламических ядер и ассоциативных зон коры, т. к. эти зоны посылают сигналы к ассоциативным ядрам. Неспецифические ядра. Афферентные сигналыполучают от других ядер таламуса по коллатералям всех сенсорных путей: от моторных центров ствола мозга, ядер мозжечка, базальных ганглиев, гиппокампа, от лобных долей. Эфферентные выходы — на другие ядра таламуса, кору больших полушарий, к другим структурам мозга. На кору оказывают модулирующее влияние, активируя ее, обеспечивают внимание.
Ассоциативные ядра
Ассоциативные ядра находятся ближе к задне-медиальной части таламуса, а также в области подушки. Особенность этих структур в том, что они не участвуют в восприятии информации, которая подходит из нижележащих образований центральной нервной системы. Эти ядра необходимы для получения уже обработанных сигналов в других ядрах таламуса или в вышележащих мозговых структурах.
Суть «ассоциативности» этих ядер в том, что к ним подходят любые сигналы, а нейроны способны их адекватно воспринять. Сигналы из этих структур поступают в области коры с соответствующим названием — ассоциативные зоны. Они расположены в височной, лобной и теменной частях коры. Благодаря поступлению этих сигналов человек способен:
Патология
Очаговые поражения Т. могут развиваться при различных патол. процессах, однако чаще всего — при нарушениях мозгового кровообращения (ишемия, кровоизлияние на почве атеросклероза и гипертонической болезни). Наиболее часты нарушения кровообращения в зоне глубоких ветвей задней мозговой артерии. При инсульте в этом бассейне в основном повреждаются вен-тролатеральные ядра Т.
или его дорсальное медиальное ядро. В Т. возможны как ишемические инфаркты, так и кровоизлияния при разрывах артерий, васкулитах, гипертонических кризах с высоким АД, при артериовенозных аневризмах. Значительно реже таламические комплексы повреждаются при инфекционных, опухолевых, дистрофических и других заболеваниях.
Воспалительные поражения Т. изредка встречаются при туберкулезе, сифилисе, сепсисе, при энцефалитах (вирусных, бактериальных). Дегенеративные процессы в структурах Т. возникают при генетических дефектах обмена веществ или при интоксикациях (экзогенных, эндогенных), а также в резидуальной стадии инфекционного или травматического поражения мозга.
Клин, симптомы при поражении Т. многообразны и зависят от функциональной роли поврежденных структур. Таламический синдром впервые подробно описан Ж. Деже-рином и Г. Русси в 1906 г. при размягчении Т. вследствие закупорки задней мозговой артерии и ее ветвей. При выключении a. thalamoge-niculata на стороне, противоположной очагу поражения в Т.
, развиваются следующие симптомы: 1) ге-мигипестезия или гемианестезия с выраженным нарушением глубокой чувствительности больше на конечностях, иногда без расстройства чувствительности на лице; 2) гиперпатия или дизестезии (см. Чувствительность, расстройства), пароксизма л ьно возникающие или постоянные жестокие боли, распространяющиеся на всю половину тела (таламический болевой синдром);
3) утрата вибрационной чувствительности; 4) преходящий гемипарез без выраженной мышечной спастичности и патол. рефлекса Бабинского; 5) атрофия мышц пострадавшей половины тела; 6) хореические и атетоид-ные движения в пальцах руки; 7) гемиатаксия; иногда гомонимная гемианопсия; 9) нотнагелевский мимический парез; 10) расстройства внимания.
При частичном поражении Т. могут преобладать отдельные группы этих симптомов. При разрушении медиальной части Т. (бассейн a. tha-lamoperforata) повреждается дентато-руброталамический путь, по к-рому в Т. поступают импульсы из мозжечка, при этом возникает гиперкинез (атетоидный, хореический) и гемиатаксия на противоположной очагу стороне.
При поражении передних отделов Т. выявляют контралатеральный мимический парез лицевой мускулатуры. При диффузном поражении Т. таламический синдром нередко сочетается с вегетативными нарушениями (цианоз, атрофия кожи и ногтей, пастозность, похолодание кожи на пораженной стороне тела), более выраженными в руке.
При этом рука приобретает характерное положение: предплечье согнуто и прони-ровано, кисть согнута в лучезапястном суставе, проксимальные фаланги пальцев согнуты, в то время как средние и дистальные фаланги разогнуты (так наз. таламическая рука). Иногда на стороне очага возникает синдром Бернара — Горнера (см. Бернара — Горнера синдром).
Диагностика таламического синдрома основывается на выявлении комплекса составляющих его симптомов. Клин, методы диагностики состоят в определении поверхностной и глубокой чувствительности, определении полей зрения (см.), силы мышц конечностей, координации движений (см.) и др. Диагностике помогает симптом Ферстера: раздражение различных рецепторов (зрительных, слуховых, вкусовых) вызывает боль и неприятное ощущение в пострадавшей половине тела.
Дифференциальный диагноз таламического синдрома Дежерина — Русси следует проводить с синдромом Геда — Холмса — односторонней аффективной дизестезией, для к-рого характерны односторонняя дизесте-зия, утрированные внешние проявления аффективных реакций (гримасничание), резкие защитные движения в ответ на незначительное физическое раздражение (напр.
, легкий укол булавкой), одностороннее расстройство вкуса и обоняния. Этот симптомокомплекс развивается при нарушении корково-таламических и корково-гипоталамических связей. Кроме того, таламический синдром следует дифференцировать с синдромом поражения внутренней капсулы, при к-ром наряду с гемианестезией (иногда в сочетании с гемианопсией) наблюдается спастическая гемиплегия. Поражение Т. можно выявить при церебральной ангиографии (см.), компьютерной томографии (см. Томография компьютерная).
Лечение таламического синдрома патогенетическое. Значительные трудности возникают при таламической гемиалгии, при к-рой обычные анальгетики мало эффективны. Рекомендуют применять седуксен, аминазин, финлепсин, тегретол и др. При гиперкинезах и неустранимых болевых синдромах применяют стереотаксические операции на ядрах Т. (см. Стереотаксическая нейрохирургия).
Прогноз таламического синдрома зависит от характера основного патол. процесса.
Библиография: Адрианов О. С. О принципах организации интегративной деятельности мозга, М., 1976; Батуев А. С. Высшие интегративные системы мозга, Д., 1981, библиогр.; ГусельниковВ. И. Электрофизиология головного мозга, М., 1976; Дуринян Р. А. Центральная структура афферентных систем, Л., 1965; он же, Корковый контроль неспецифических систем мозга, М., 1975; Кесарев В. С.
Количественная архитектоника мозга человека, Вестн. АМН СССР, № 12, с. 29, 1978; Клиническая нейрофизиология, под ред. Н. П. Бехтерева, с. 49, Л., 1972; Кроль М. Б. и Федорова Е. А. Основные невропатологические синдромы, М., 1966; Курепина М. М. Мозг животных, М., 1981;Общая и частная физиология нервной системы, под ред. П. Г. Костюка, с. 313, Л., 1969; Серков Ф. Н. и Казаков В. Н.
Нейрофизиология таламуса, Киев, 1980, библиогр.; Сосудистые заболевания нервной системы, под ред. Е. В. Шмидта, с. 56, 330, М., 1975; Экспериментальная психология, под ред. С. С. Стивенса, пер. с англ., с. 174, М., 1960; В a r r a q u е r-В о г- d a s L. а. о. Thalamic hemorrhage, Stroke, v. 12, p. 524, 1981; Brow n J. W.
Thalamic mechanisms in language, Handb. behavioral neurobiol., ed. by F. A. King, v. 2, p. 215, N. Y, —L., 1979; DantzerR. etDela-c о u r J. Modification d’un phenomene de suppression conditionnee par une leson thalamique, Physiol. Behav., t. 8, p. 997, 1972; Dejerine J. a. Roussy G. Le syndrome thalamique, Rev. neurol. (Paris), t. 14, p. 521, 1906;
Einfiihrung in die stereotaktischen Operationen mit einem Atlas des menschlichen Gehirns, hrsg. v. G. Schaltenbrand a. P. Bailey, Bd 1, S. 230, 1959, Bibliogr.; Graeber R. C. a. Ebbesson S.O. Retinal projections in the lemon shark (Negaprion previrostris), Brain Behav. Evolut., v. 5, p. 461, 1972; G r a y b i e 1 A. M.
Some fiber pathways related to the posterior thalamic region in the cat, ibid., v. 6, p. 363, 1972; Henderson V. W., Alexander M. P. a. N a e s e г M. A. Right thalamic injury, impaired visuospatial perception, and alexia, Neurology, v. 32, p. 235, 1982, bibliogr.; K w a k R., K a d о у a S. a. Suzuki T.
Factors affecting the prognosis in thalamic hemorrhage, Stroke, v. 14, p. 493, 1983; Lapresle J. a. Ha-g и e n а и M. Anatomico-chemical correlation in focal thalamic lesions, Z. Neurol., v. 205, p. 29, 1973; L e j e u n e H. Lesions thalamiques medianes et effets differentiels dans les apprentissages operants, Physiol, a. Behav., t. 18, p. 349, 1977;
Van В u-r e n J. М. а. В o r k e R. C. Variations and connections of the human thalamus, pt 1 — 2, N. Y. a. o., 1972; V e 1 a s с o F. a. Velasco M. A reticulothalamic system mediating proprioceptive attention and tremor in man, Neurosurgery, v. 4, p. 30, 1979; Villablanca J. a. Sali-nas-Zeballos М. E. Sleep-wakeful-ness, EEG and behavioral studies of chronic cats without the thalamus, Arch. ital. Biol., v. 110, p. 383, 1972.
Д. К. Богородинский, А. А. Скоромец; О. С. Адрианов (физ.), В. С. Кесарев (ан., сравнительная анатомия).
Неспецифические ядра
Неспецифическими эта группа ядер называется потому, что получает информацию практически от всех структур центральной нервной системы:
Импульс от неспецифических ядер также идет ко всем областям коры головного мозга. Такая выборочность, как в случае с ассоциативными и специфическими ядрами, здесь отсутствует.
Так как именно эта группа ядер имеет наибольшее количество связей, считается, что благодаря ей обеспечивается слаженная, координированная работа всех участков головного мозга.
Отдельно о зрительном бугре
Ранее считалось, что таламус обрабатывает только зрительные импульсы, тогда орган и получил название — зрительные бугры. Сейчас это название считается устаревшим, так как орган обрабатывает практически весь спектр афферентных систем (кроме обоняния).
Система, которая обеспечивает зрительное восприятие – одна из самых интересных. Основной внешний орган зрения – глаз – рецептор, который имеет сетчатку и оснащен особенными клетками (колбочки, палочки), которые трансформируют световой пучок и электрический сигнал. Электрический сигнал, в свою очередь, проходя по нервным клеткам, попадает в латеральный центр таламуса, который отправляет обработанный сигнал в центральный отдел коры головного мозга. Тут происходит окончательный анализ сигнала, благодаря чему формируется увиденное, то есть – картинка.
Метаталамус
Отдельно выделяют группу ядер зрительного бугра под названием метаталамус. Данная структура состоит из медиального и латерального коленчатых тел.
Медиальное коленчатое тело получает информацию о слухе. Из нижележащих отделов мозга информация поступает через верхние горбики среднего мозга, а сверху структура получает импульс из слуховой области коры.
Латеральное коленчатое тело относится к зрительной системе. Чувствительная информация к ядрам этой группы поступает от сетчатки глаза через зрительные нервы и зрительный тракт. Обработанная в таламусе информация далее идет к затылочной области коры, где находится первичный центр зрения.
Функции таламуса
Как происходит обработка поступающей с периферии чувствительной информации, которая далее передаются в кору переднего мозга? Это и есть основная роль зрительного бугра.
Благодаря этой функции при повреждении коры возможно восстановление чувствительности посредством таламуса. Таким образом, возможна репарация болевого, температурного чувства, а также грубого осязания.
Еще одна важная функция таламуса — это координация движений и чувствительности, то есть сенсорной и моторной информации. Это обусловлено тем, что в таламус поступают не только сенсорные импульсы. Также к нему идут импульсы от мозжечка, ганглиев экстрапирамидной системы, коры большого мозга. А эти структуры, как известно, принимают участие в осуществлении движений.
Также зрительный бугор участвует в поддержании сознательной активности, регуляции сна и бодрствования. Эта функция осуществляется за счет наличия связей с голубым пятном ствола мозга и гипоталамусом.
Таламус, как предполагаемый биомаркер психических расстройств
Особенности таламической морфологии при различных нейропсихиатрических расстройствах могут предоставить важную информацию о начале, прогрессировании и исходе расстройство , а также ответе на лечение. Созревание таламуса и коры тесно связано, так что таламическая аномалия в раннем периоде развития нервной системы может ухудшить нормальное развитие коры (и наоборот), и, что не удивительно, патология зрительного бугра вовлечена в нейробиологию шизофрении. Таламус (от греческого слова thalamos, используемый для обозначения самой внутренней комнаты, кладовой ) можно разделить на дорсальные и вентральные отделы: дорзальный таламус состоит из ядер, имеющих взаимные связи с корой головного мозга и полосатым телом , тогда как вентральный таламус обычно не посылает свои проекции в кору. У людей дорсальный таламус представляет собой парную структуру со стратегическим центральным расположением между другими подкорковыми структурами и корой. Два отдельных дорзального таламуса расположены у основания каждого полушария головного мозга по обе стороны от третьего желудочка соответственно. Дорзальный таламус имеет передние, медиальные и боковые подразделения, определяемые изогнутым листом миелинизированных волокон, называемым внутренней медуллярной пластинкой. Эти подразделения состоят из различных ядер таламуса, а их разделение основанно на цитоархитектуре, структурах связности и функциональности, при этом эти ядра обладают многими функциями.
Существует два типа нейронов в дорзальных таламических ядрах, которые можно отличить по их морфологии и хемоархитектуре: локально действующие гамкергические интернейроны и глутаматергические ретрансляционные клетки, выступающие за пределы таламуса. Физиологические свойства нейлонов ретрансляции таламуса показывают, что они могут подвергаться двум основным типам режимов отклика, которые будут определять характер сообщения, передаваемого в кору: считается, что режим импульсного ответа используется для обнаружения сигнала, тогда как режим тонического ответа который, как полагают, используется таламическими ретрансляционными ячейками для более точного анализа сигналов. Было высказано предположение, что нарушение процессов модуляции (т. е. переход от анализа сигнала к детектированию сигнала) может привести к аберрантной значимости, например, наблюдаемой при шизофрении.
Считается, что ядра дорзального таламуса получают два типа афферентных волокон, которые классифицируются как модуляторы, независимо от их происхождения (т. е. коркового или подкоркового), и которые могут быть определены на основе их синаптической морфологии и постсинаптических действий. В этом смысле драйверами являются по существу те афференты к таламусу, которые переносят сообщение, передаваемое таламокортикальными клетками, тогда как модуляторы — это афференты к таламусу, которые влияют на то, как это сообщение передается этими таламокортикальными клетками. Таламические ядра можно классифицировать по афферентам, которые они получают. Первичные (или первичные) ядра получают свои драйверы из периферийной или подкорковой структуры и получают модуляторы из пирамидальных клеток в слое VI ипсилатеральной коры. В этой схеме визуальные, соматосенсорные и слуховые афференты посылают периферическую сенсорную информацию в ядра первого порядка таламуса. С другой стороны, ядра высшего порядка (или ассоциации) получают свои драйверы из пирамидальных клеток ,находящихся в слое V ипсилатеральной коры. В этой схеме реле более высокого порядка передают сообщения от одной области коры до другой, например, относительно выхода на моторные или премоторные центры из кортикальной области. Подобно тому, как сенсорная информация проходит через ядра первого порядка по пути к коре, так и кортико — кортикальная информация проходит через ядра более высокого порядка (с прямыми кортико — кортикальными путями, обеспечивающими некоторую другую функцию, например, модуляторную); что бросает вызов традиционному представлению о кортико — кортикальной передаче сигналов. Что касается режима реакции ретрансляционных ячеек в ядрах более высокого порядка, считается, что импульс ( всплеск ) и тонические моды используются аналогичным образом: режим всплеска может быть задействован для указания сдвига в структуре выхода в транстохостической кортико — кортикальной связи (от одной кортикальной область к другой), и тонизирующий режим затем включается ячейками реле более высокого порядка для более надежной передачи информации в другую область коры. Следует отметить, что исследования показывают, что в ядрах более высокого порядка больше разрывов, чем в реле первого порядка. Несмотря на ограниченность, имеющиеся данные свидетельствуют о том, что драйверы для реле более высокого порядка исходят из нескольких различных областей коры, что предполагает, что реле, вероятно, будет представлять несколько различных функций. По меньшей мере половина таламуса вовлечена в транстохастическую кортико — кортикальную коммуникацию, это подразумевает, что нарушение такого типа сигнализации может влиять на более высокие кортикальные функции. Функция таламо — кортикальных входов в кору (как в первом, так и в более высоком порядке) может заключаться в том, чтобы обновить кору в последнем при моторных командах.
Традиционно при визуализации отображаются многие из афферентных и эфферентных связей таламуса, в том числе афферентные связи для ядер первого порядка, а также топографически организованные пути таламо — кортикального и кортико — таламического типа первого и более высокого порядка. Транс-синаптическая нейродегенерация была отмечена, как механизм нейродегенеративного заболевания, при котором деафферентация от кортикальных нейронов может привести к синаптической дисфункции и, следовательно, распространению патологии нейронов по уязвимым нейронным сетям. Это может быть одним из механизмов, который приводит к подкорковой патологии, такой как потеря нейронов при нейродегенеративных заболеваниях, о чем свидетельствует поражение полосатого тела при болезни Хантингтона (HD), лобно-височной деменции (FTD) и болезни Альцгеймера (AD). Учитывая топографическую связь таламуса с корой, можно было бы предсказать, что таламус может служить картой структурных изменений кортикальных афферентных путей при различных нейродегенеративных заболеваниях.
С точки зрения нейронной цепи существует ряд четко определенных анатомических петель, связанных с таламусом. Например, считается, что линии фронтальной — стриатной таламо — кортикальной петли образуют основную сеть, благодаря которой двигательная активность и поведение опосредуются, и могут объяснить сходство поведенческих изменений при лобных кортикальных и подкорковых расстройств. Таламус является критическим центром в этих сетях. Предполагается, что кортико-подкорковые поражения при процессах нейродегенеративных заболеваний могут разъединять эти схемы, и это лежит в основе проявленной нейропсихиатрической дисфункции, такой как когнитивные и поведенческие нарушения, наблюдаемые, например, при FTLD. Аналогичным образом, дисфункция мозжечково-локулярных путей также связана с нейропсихиатрическим заболеванием. Нарушение нейронных цепей, связывающих кору, таламус и мозжечок, может играть в патогенезе шизофрении значимую роль , полагая, что нарушение этой схемы может зависеть от эмоциональной и когнитивной дисфункции.
Исключая большинство обонятельных входов, все сенсорные тракты на пути к коре головного мозга сначала проходят через дорзальный таламус. Считалось, что функция дорзального таламуса не ограничивается передачей информации, и действительно она была описана как «шлюз» к коре. С точки зрения текущей концептуализации таламической функции считается, что таламическое реле функционирует как блокирующий вентиль (а не интегратор из нескольких сообщений) с его ячейками реле компонентов, передающими сообщение от их афферентного движения (будь то кортикальный или подкорковый ) к коре; сигнализация происходит либо в импульсном, либо в тоническом режимах, хотя и с небольшими изменениями. Правда не совсем ясно, какое преимущество сохраняется, когда отправляется информацию через таламус, прежде чем она достигнет области коры. При концептуализации роли таламуса при нейропсихиатрических расстройствах будет полезно рассмотреть отказ нервных схем, связанных с таламусом, а не нейропсихиатрические симптомы, возникающие непосредственно из самого таламуса. Роль таламической патологии в состоянии болезни тогда можно было бы считать неудачей ее релейных функций или ненормальности стробирования, которая действует на сообщения, которые она получает.
При шизофрении отмечатеся объемный дефицит таламуса относительно размера мозга , и при этом показано, что таламический размер отрицательно коррелирует с серьезностью симптомов шизофрении. Таламический дефицит объема был зарегистрирован при психозах первого эпизода, у пациентов с плохими лекарственными препаратами для лечения шизофрении, а также у тех пациентов, которые принимали антипсихотические препараты, и имеются противоречивые данные о том, увеличивается ли объем таламика при лечении антипсихотическими препаратами.