Строение и функции
Физиология нервной системы является сложной составной структурой.
Нейрон считается основной структурной и функциональной единицей нервной системы. Его отростки формируют волокна, которые возбуждаются при воздействии и передают импульс. Импульсы достигают центров, где подвергаются анализу. Проанализировав полученный сигнал, мозг передает необходимую реакцию на раздражитель соответствующим органам или частям тела. Нервная система человека кратко описывается следующими функциями:
- обеспечение рефлексов;
- регуляция внутренних органов;
- обеспечение взаимодействия организма с внешней средой, путем приспособления тела к изменяющимся внешним условиям и раздражителям;
- взаимодействие всех органов.
Значение нервной системы заключается в обеспечении жизнедеятельности всех частей организма, а также взаимодействии человека с окружающим миром. Строение и функции нервной системы изучаются неврологией.
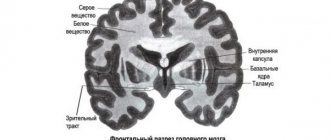
Вещество мозга
- Серое вещество образовано телами нейронов и короткими отростками – дендритами
- Белое вещество образованно длинными отростками нейронов – аксонами.
- Нервная регуляция осуществляется за счет нервного импульса.
- Нервный импульс – это электрическая волна, бегущая по нервному волокну.
- Преобразует раздражение в нервный импульс рецептор.
- Деятельность нервной системы носит рефлекторный характер.
- Рефлекс – это ответная реакция организма на раздражение, происходящая при участии нервной системы.
- Нервный импульс, возникший при раздражении, проходит определенный путь, называемый рефлекторной дугой.
- Рефлекторная дуга — путь, по которому проходят нервные импульсы (нервное возбуждение) от рецепторов к исполнительному (рабочему) органу при осуществлении рефлекса; включает пять отделов:
Структура ЦНС
Анатомия центральной нервной системы (ЦНС) является скоплением нейронных клеток и нейронных отростков спинномозгового отдела и головного мозга. Нейрон – это единица нервной системы.
Функция ЦНС – это обеспечение рефлекторной деятельности и обработка импульсов, поступающих от ПНС.
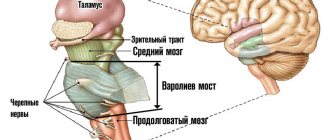
Анатомия центральной нервной системы, основным узлом которой является головной мозг, представляет собой сложную структуру из разветвленных волокон.
В больших полушариях сосредоточены высшие нервные центры. Это – сознание человека, его личность, его интеллектуальные способности и речь. Основная функция мозжечка – это обеспечение координации движений. Ствол мозга неразрывно связан с полушариями и мозжечком. В этом отделе находятся основные узлы двигательных и чувствительных проводящих путей, благодаря чему обеспечиваются такие жизненно важные функции организма, как регуляция кровообращения и обеспечение дыхания. Спинной мозг является распределительной структурой ЦНС, он обеспечивает разветвление волокон, образующих ПНС.
Спинномозговой узел (ганглий) является местом сосредоточения чувствительных клеток. С помощью спинномозгового ганглия осуществляется деятельность вегетативного отдела периферической нервной системы. Ганглии или нервные узлы в нервной системе человека относят к ПНС, они выполняют функцию анализаторов. Ганглии не относятся к центральной нервной системе человека.
Особенности строения ПНС
Благодаря ПНС происходит регулирование деятельности всего организма человека. ПНС состоит из черепных и спинномозговых нейронов и волокон, образующих ганглии.
У периферической нервной системы человека строение и функции очень сложные, поэтому любое малейшее повреждение, например, повреждение сосудов на ногах, может вызвать серьезные нарушения ее работы. Благодаря ПНС осуществляется контроль за всеми частями организма и обеспечивается жизнедеятельность всех органов. Значение этой нервной системы для организма переоценить невозможно.
ПНС делится на два подразделения – это соматическая и вегетативная системы ПНС.
Соматическая нервная система выполняет двойную работу – сбор информации от органов чувств, и дальнейшая передача этих данных в ЦНС, а также обеспечение двигательной активности организма, путем передачи импульсов от ЦНС в мышцы. Таким образом, именно нервная система соматическая является инструментом взаимодействия человека с окружающим миром, так как она обрабатывает сигналы, получаемые от органов зрения, слуха и вкусовых рецепторов.
Вегетативная нервная система обеспечивает выполнение функций всех органов. Она контролирует сердцебиение, кровоснабжение, дыхательную деятельность. В ее составе – только двигательные нервы, регулирующие сокращение мышц.
Для обеспечения сердцебиения и кровоснабжения не требуются усилия самого человека – этим управляет именно вегетативная часть ПНС. Принципы строения и функции ПНС изучаются в неврологии.
Строение нервной системы
Возбудимость, раздражимость и проводимость характеризуются как функции времени, то есть это – процесс, возникающий от раздражения до появления ответной реакции органа. Распространение нервного импульса в нервном волокне происходит за счет перехода локальных очагов возбуждения на соседние неактивные области нервного волокна. Нервная система человека обладает свойством трансформации и генерации энергий внешней и внутренней среды и преобразования их в нервный процесс.
Строение нервной системы человека: 1- плечевое сплетение; 2- кожно-мышечный нерв; 3- лучевой нерв; 4- срединный нерв; 5- подвздошно-подчревный нерв; 6- бедренно-половой нерв; 7- запирающий нерв; 8- локтевой нерв; 9- общий малоберцовый нерв; 10- глубокий малоберцовый нерв; 11- поверхностный нерв; 12- мозг; 13- мозжечок; 14- спинной мозг; 15- межреберные нервы; 16- подреберный нерв; 17- поясничное сплетение; 18- крестцовое сплетение; 19- бедренный нерв; 20- половой нерв; 21- седалищный нерв; 22- мышечные ветви бедренных нервов; 23- подкожный нерв; 24- большеберцовый нерв
Нервная система функционирует как единое целое с органами чувств и управляется головным мозгом. Самая крупная часть последнего называется большими полушариями (в затылочной области черепа находятся два более мелких полушария мозжечка). Головной мозг соединяется со спинным. Правое и левое большие полушария соединены между собой компактным пучком нервных волокон, называемых мозолистым телом.
Спинной мозг – основной нервный ствол тела – проходит через канал, образованный отверстиями позвонков, и тянется от головного мозга до крестцового отдела позвоночника. С каждой стороны спинного мозга симметрично отходят нервы к различным частям тела. Осязание в общих чертах обеспечивается определенными нервными волокнами, бесчисленные окончания которых находятся в коже.
Отделы ПНС
ПНС также состоит из афферентной нервной системы и эфферентного отдела.
Афферентный отдел представляет собой совокупность сенсорных волокон, которые обрабатывают информацию от рецепторов и передают ее в головной мозг. Работа этого отдела начинается тогда, когда рецептор раздражается из-за какого-либо воздействия.
Эфферентная система отличается тем, что обрабатывает импульсы, передающиеся от головного мозга к эффекторам, то есть мышцам и железам.
Одна из важных частей вегетативного отдела ПНС – это энтеральная нервная система. Энтеральная нервная система формируется из волокон, расположенных в ЖКТ и мочевыделительных путях. Энтеральная нервная система обеспечивает моторику тонкой и толстой кишки. Этот отдел также регулирует секрет, выделяемый в ЖКТ, и обеспечивает местное кровоснабжение.
Вегетатика
Классификация нервных систем людей по их строению – не единственный способ разделения ее отделов. Колоссальное значение имеет вегетативная нервная система, осуществляющая непосредственное управление органами всех систем. Человек не может осознанно управлять деятельностью вегетатики, но информация о том, как она работает, помогает иногда скорректировать самочувствие при вегетативных нарушениях, например, при распространенном заболевании – ВСД (вегетососудистая дистония).
Деятельность вегетативной нервной системы осуществляется за счет двух отделов-антагонистов: симпатического и парасимпатического. То есть, когда активизируется симпатический отдел, деятельность парасимпатического автоматически прекращается.
Внутриутробное развитие ЦНС
Значение нервной системы заключается в обеспечении работы внутренних органов, интеллектуальной функции, моторике, чувствительности и рефлекторной деятельности. ЦНС ребенка развивается не только во внутриутробный период, но и на протяжение первого года жизни. Онтогенез нервной системы начинается с первой недели после зачатия.
Основа для развития головного мозга формируется уже на третьей неделе после зачатия. Основные функциональные узлы обозначаются к третьему месяцу беременности. К этому сроку уже сформированы полушария, ствол и спинной мозг. К шестому месяцу высшие отделы мозга уже развиты лучше, чем спинальный отдел.
К моменту появления малыша на свет, наиболее развитым оказывается головной мозг. Размеры мозга у новорожденного составляют примерно восьмую часть веса ребенка и колеблются в пределах 400 г.
Деятельность ЦНС и ПНС сильно понижена в первые несколько дней после рождения. Это может заключаться в обилии новых раздражающих факторов для малыша. Так проявляется пластичность нервной системы, то есть способностью этой структуры перестраиваться. Как правило, повышение возбудимости происходит постепенно, начиная с первых семи дней жизни. Пластичность нервной системы с возрастом ухудшается.
Типы ЦНС
В центрах, расположенных в коре мозга, одновременно взаимодействуют два процесса – торможение и возбуждение. Скорость смены этих состояний определяет типы нервной системы. В то время как возбужден один участок центра ЦНС, другой замедляется. Этим обусловлены особенности интеллектуальной деятельности, такие как внимание, память, сосредоточенность.
Типы нервной системы описывают отличия между скоростью процессов торможения и возбуждения ЦНС у разных людей.
Люди могут отличаться по характеру и темпераменту, в зависимости от особенностей процессов в ЦНС. К ее особенностям относят скорость переключения нейронов с процесса торможения на процесс возбуждения, и наоборот.
Типы нервной системы делятся на четыре вида.
- Слабый тип, или меланхолик, считают наиболее предрасположенным к возникновению неврологических и психоэмоциональных расстройств. Он отличается медленными процессами возбуждения и торможения. Сильный и неуравновешенный тип – это холерик. Этот тип отличается преобладанием процессов возбуждения над процессами торможения.
- Сильный и подвижный – это тип сангвиника. Все процессы, проистекающие в коре головного мозга сильны и активны. Сильный, но инертный, или флегматический тип, отличается низкой скоростью переключения нервных процессов.
Типы нервной системой взаимосвязаны с темпераментами, но эти понятия следует различать, ведь темперамент характеризует набор психоэмоциональных качеств, а тип ЦНС описывает физиологические особенности процессов, происходящих в ЦНС.
Защита ЦНС
Анатомия нервной системы очень сложная. ЦНС и ПНС страдают из-за воздействия стресса, перенапряжения и недостатка питания. Для нормального функционирования ЦНС необходимы витамины, аминокислоты и минералы. Аминокислоты принимают участие в работе мозга и являются строительным материалом для нейронов. Разобравшись, зачем и для чего нужны витамины и аминокислоты, становится ясно, как важно обеспечить организм необходимым количеством этих веществ. Особенно для человека важны глютаминовая кислота, глицин и тирозин. Схема приема витаминно-минеральных комплексов для профилактики заболеваний ЦНС и ПНС подбирается индивидуально лечащим врачом.
Повреждения пучков нервных волокон, врожденные патологии и аномалии развития мозга, а также действие инфекций и вирусов – все это приводит к нарушению работы ЦНС и ПНС и развитию различных патологических состояний. Такие патологии могут вызвать ряд очень опасных заболеваний — обездвиживание, парез, атрофия мышц, энцефалит и многое другое.
Злокачественные новообразования в головном или спинном мозге приводят к ряду неврологических нарушений. При подозрениях на онкологическое заболевания ЦНС назначается анализ — гистология пораженных отделов, то есть обследование состава ткани. Нейрон как часть клетки также может мутировать. Такие мутации позволяет выявить гистология. Гистологический анализ проводится по показаниям врача и заключается в сборе пораженной ткани и ее дальнейшем изучении. При доброкачественных образования также проводится гистология.
В теле человека находится множество нервных окончаний, повреждение которых может вызвать ряд проблем. Повреждение зачастую приводит к нарушению подвижности части тела. Например, повреждение руки может привести к боли на пальцах рук и нарушению их движения. Остеохондроз позвоночника спровоцировать возникновение болей на стопе из-за того, что раздраженный или передавленный нерв посылает болевые импульсы рецепторам. Если болит ступня, люди часто ищут причину в долгой ходьбе или травме, но болевой синдром может быть спровоцирован повреждением в позвоночнике.
При подозрении на повреждение ПНС, а также при любых сопутствующих проблемах необходимо пройти осмотр у специалиста.
Будь умным!
Классическое исследование параметров и механизма ПД проделано в работах Ходжкина и Хаксли на гигантском аксоне кальмара с отведением внутриклеточного потенциала и внутриклеточным раздражением. В это нервное волокно (диаметр d волокна 0,5-1 мм) вводили два тончайших коаксиальных проволочных электрода, один из которых регистрировал электрический потенциал относительно внешнего электрода, другой — был раздражающим: через него в волокно подавались толчки тока того или иного направления (рис.3.3 ). При подаче короткого и слабого толчка выходящего тока (направление его определяется для положительных зарядов) внутриклеточный электрод регистрировал кратковременное падение мембранного потенциала (МП), по форме и по силе соответствующего толчку тока, но со сглаженными передним и задним фронтами (сглаженность фронтов определяется емкостью мембраны). Это так называемый электротонический потенциал (ЭП) При подаче несколько более сильного толчка выходящего тока к электротоническому потенциалу присоединяется дополнительная деполяризация, называемая подпо-роговым или «локальным» ответом (ЛО). Локальным он называется потому, что и в этих и в естественных условиях не распространяется. А если стимул еще усиливается и достигается критический уровень деполяризации (КУД), то возникает потенциал действия (рис. ). При толчке входящего тока любой силы получается только электротонический потенциал.
В потенциале действия (ПД) различают пик (спайк) и следовые потенциалы.
Рис.3.3. Схемы методик, применяемых на различных нервных волокнах для исследования их электрогенеза
a — внутриклеточное раздражение и отведение потенциалов гигантского аксона кальмара при коаксиальном введении электродов .
б — раздражение и отведение потенциалов от одиночного перехвата Ранвье, изолированного двумя воздушными промежутками («мостиками»).
Пик ПД представляет собой кратковременную инверсию внутриклеточного потенциала. Он имеет очень быструю S-образную восходящую фазу и несколько более медленный спад. Общая длительность пика в данном объекте приблизительно равна 3 мс. Амплитуда пика ∼110 мВ, т. е. она превышает МПП (-70 мВ) на 40 мВ. Эта разница называется овершутом («перелетом»). Вслед за пиком ПД регистрируются значительно более слабые и длительные отрицательный и далее положительный следовой потенциалы (рис.3.4 ). ПД имеет стандартные параметры, не зависящие от силы вызвавшего ПД стимула.
Рис.3.4. Схема регистрации основных электрофизиологических феноменов в нервном волокне.
МП — мембранный потенциал покоя, ПД — потенциал действия, ЛО — локальный ответ, КУД — критический уровень деполяризации, СН — следовая негативность, СП — следовая позитивность, АЭТ — анэлектротонический потенциал, КЭТ — катэлектротонический потенциал.
При раздражении гигантского нервного волокна выходящим током через коаксиальный электрод все точки этого нервного проводника раздражаются практически равномерно, и в них одновременно возникает и развивается ПД. Такой ПД, по существу, не распространяется и называется «мембранным». В естественных условиях ПД лишь возникает локально, а затем распространяется (проводится) вдоль волокна. Это распространяющийся ПД. Мембранный ПД несколько проще распространяющегося, что делает его более удобным для анализа. Заметим, что в миелинизированных волокнах мембранный ПД получают при работе на изолированном одиночном перехвате Ранвье.
Какова природа потенциала действия?
Прежде всего ПД — это электрический феномен, возникающий на плазматической мембране и в связи с ее активностью. Бейкер, Шоу, Ходжкин показали, что практически нормальный ПД возникает и в перфузируемом гигантском аксоне, лишенном аксоплазмы, при электрической стимуляции его мембраны.
Причиной развития ПД является вызываемое электростимуляцией, точнее критической деполяризацией мембраны, открытие потенциалозависимых каналов в мембране.
Уже в 1939 г. Кол и Кэртис нашли, что при развитии ПД резко снижается поперечный импеданс аксона кальмара за счет падения его RM (с 1000 до 25 Ом • см2) при неизменной См. Как показали последующие исследования это связано с открытием потенциалозависимых натриевых и калиевых каналов. Заметим, что участвующие в формировании МПП каналы утечки потенциалонезависимы. Открытие потенциалозависимых каналов приводит к пассивному движению соответствующих ионов по их электрохимическим градиентам. Причем вход ионов натрия в волокно обеспечивает восходящую фазу пика ПД, деполяризацию и инверсию потенциала на мембране, а несколько запаздывающий выход ионов калия участвует в создании нисходящей фазы пика, реполяризации. При развитии пика ПД отношение Рк: PNА: PCI становится равным 1 : 20 : 0,45 (в покое оно = 1 : 0,04:0,45). Связь развития пика ПД с током Na+ доказывается следующими обстоятельствами: 1) прямой зависимостью амплитуды ПД от электрохимического градиента Na+ на мембране, 2) достоверным переходом меченого 24Na из среды в волокно при его возбуждении, причем в количестве, пропорциональном числу ПД.
Связь нисходящей фазы ПД с током К+ доказывается зависимостью хода этой фазы от электрохимического градиента К+ на мембране.
Подробный анализ изменений мембранной проницаемости для ионов Na+ и К+, основанный на измерениях токов этих ионов, стал возможным благодаря использованию методики фиксации («кламп») электрического потенциала мембраны, примененной впервые Колом, Мармонтом (1949). Заметим, что мембранные токи (при данных концентрационных градиентах) зависят и от ионной проницаемости) и от мембранного потенциала .Поэтому мембранные токи могут точно характеризовать изменения Р только при МП = = const.
Методика фиксации мембранного потенциала состоит в следующем. Мембрана гигантского волокна (или какой-либо иной клетки) с помощью внутриклеточного и наружного электродов соединяется с электронной схемой. Основной частью этой схемы является дифференциальный усилитель, сравнивающий МП с задаваемым от постороннего источника потенциалом Е. Ток (I) на выходе усилителя определяется по направлению и по величине разностью Е-МП, подаваемой на вход. Этот ток (/) протекает через мембрану и создает на ней дополнительный скачок потенциала, уменьшающий абсолютную величину раз-ности Е-МП. При большом коэффициенте усиления дифференциального усилителя и малой величине R в цепи, подводящей ток, компенсирующий скачок потенциала на мембране может быть весьма приближен по величине к разности Е-МП и таким образом, в итоге МП может стать приблизительно равным Е.
В этой ситуации при достаточном быстродействии системы (при τ<20 мкс) любое, даже слабое, случайное смещение МП немедленно компенсируется. Таким образом, МП фиксируется на величине Е и вместе с тем управляется путем изменения этой величины Е.
Методика фиксации потенциала на мембране исключает любые смещения МП, в том числе потенциалы действия. Но она позволяет регистрировать и исследовать трансмембранные токи, возникающие при искусственном изменении МП (Е). Если фиксированный МП равен потенциалу покоя, то трансмембранного тока практически нет. Если МП скачкообразно повышают, то возникают только направленный внутрь очень краткий емкостный ток и за ним постоянный ток утечки полиионной, в основном калиевой природы. Но если МП скачком снижают, то вслед за током разряда мембранной емкости, на фоне тока утечки, направленного наружу, возникает дополнительно-краткий (так называемый «быстрый») ток внутрь и за ним более длительный (так называемый «медленный» ток) наружу. Последние два тока-это ионные токи, текущие через натриевые и калиевые каналы, открываемые деполяризацией. Если устранить натриевый градиент на мембране путем замены части Na+ на холин, или довести МП до величины, равной ENа, то картина преобразуется: исчезает быстрый, внутрь направленный ток, но зато выявляется в неосложненном виде медленный ток, направленный наружу. Того же можно добиться специфической блокадой натриевых каналов, применив тетродотоксин (ТТХ). Таким образом, быстрый внутрь направленный ток — это натриевый ток. Картина его развития может быть получена путем геометрического вычитания медленного ионного тока из суммарного. Медленный (иначе «задержанный») ионный ток — это калиевый ток, его сила пропорциональна калиевому электрохимическому градиенту и PK. Сейчас установлено, что этот ток устраняется блокадой калиевых каналов тетраэтиламмонием (ТЭА), прилагаемым снаружи, и ионами цезия (Cs+) изнутри.
Быстрый входящий INa и более медленный выходящий Iк начинаются в общем одновременно, но INa быстрее развивается, достигает своего «потолка». Заметим, что если Е (МП) возвращается к величине, характерной для покоя, то натриевый ток исчезает примерно в 10 раз быстрее, чем калиевый. Иначе говоря, в массе натриевые каналы мембраны и активируются (деполяризацией), и деактивируются (реполяризацией) быстрее, чем калиевые каналы. Заметим, что активация каналов определяется открытием их активационных ворот, а деактивизация закрытием этих же ворот (рис.3.5 ).
Возвращаясь к картине развития INa и IK при длительной деполяризации (рис. ) в условиях клампа, обратим внимание на то, что INa уже через 5 мсек исчезает, несмотря на деполяризованное состояние мембраны (IK при этом сохраняется). Этот феномен называют инактивацией натриевых каналов. Его связывают с закрытием специальных инактивационных ворот в натриевых каналах. Схема срабатывания активационных ворот натриевых каналов (m-ворот), а также их инактивационных ворот (h-ворот) при разных МП дана на рис. . Инактивация натриевых каналов развивается из-за деполяризации, т. е., по-видимому, по той же причине, что и активация, но позже, что и делает возможным INa (а значит, и ПД в естественных условиях). Инактивация РNа — это очень важный механизм, способствующий прекращению пика ПД, лежащий в основе невозбудимости — рефрактерности после возбуждения (рефрактерная фаза), катодической депрессии и парабиотического состояния нервной (мышечной) клетки, связанного с деполяризацией. Заметим, что инактивация Na-каналов после реполяризации мембраны проходит. Инактивация калиевых каналов в данном объекте развивается очень медленно.
Рис.3.5. Схема работы ворот потенциалозависимых натриевых каналов нервной мембраны.
а — зависимость процента открытых инактивационных (h) и активационных (τ) ворот от мембранного потенциала, б — схематическое изображение положений τ и h — ворот при покое (1), развитии пика ПД (2) и в рефрактерной фазе (3).
Теперь обратимся к характеристике натуральных ионных токов, лежащих в основе пика ПД. Эти ионные токи в общем сходны с получаемыми при критической деполяризации в методике клампа, но развиваются и прекращаются они гораздо быстрее. Дело в том, что развитие ПД — это сильное изменение МП, которое влияет на трансмембранные ионные токи. В то же время эти ионные токи влияют на МП.
В процессе развития ПД действуют многие факторы , связанные прямыми и обратными связями.
Например, на восходящей фазе ПД действует система факторов с положительной обратной связью:
Здесь деполяризация увеличивает РNа , а последняя порождает INa, усиливающий деполяризацию (пока и поскольку INa больше Iк+I утечки). В силу этих отношений INa в ПД и восходящая фаза самого ПД развивается быстро, с ускорением, а амплитуда ПД всегда достигает некоторого максимума, более или менее приближенного к ENa (в зависимости от степени компенсации натриевого тока токами К+ и утечки). На спаде ПД при реполяризации для INa действует та же система факторов, но с обратными знаками эффектов. А для калиевого тока действует система факторов с отрицательной обратной связью:
В итоге спад Iк несколько затягивается.
Если скрупулезно учесть все эти факторы и их взаимодействие, то можно из величин токов, получаемых при фиксации мембранного потенциала на разных уровнях, рассчитать форму нормального ПД, что и сделали А. Хаксли и А. Ходжкин. Было показано совпадение расчетных параметров ПД и параметров ПД, регистрируемых в опыте.
Расход ионов на пик одного проводящегося ПД даже в гигантском аксоне кальмара очень мал. Например, расход внутриаксонального К при этом приблизительно равен 1-й миллионной доле внутреннего калиевого запаса. Таким образом, пик ПД — весьма экономичный сигнал, практически не нарушающий ионных градиентов на мембране, энергией которых он питается. Ионные градиенты на мембране — это «пружина», энергии которой может хватить на 5 • 105 ПД без подзарядки. Но для длительной работы волокна ионные градиенты нужно восстанавливать, что и обеспечивает работа натрий-калиевого насоса мембраны.
Несколько слов о механизме подпорогового локального ответа (ЛО) в нервном волокне. Этот ответ имеет в общем тот же механизм, что и ПД. Его восходящая фаза определяется входящим током Na+, а нисходящая — выходящим калиевым током. Но амплитуда ЛО (и его натриевый ток Na) пропорциональна силе подпорогового раздражения, а не стандартна, как у ПД, т. е. не подчиняется правилу «все или ничего». ЛО не вызывает абсолютной рефрактерности, хотя обнаруживает относительную рефрактерность из-за инактивации части натриевых каналов.
Необходимо остановиться на некоторых особенностях потенциалов действия в телах нейронов моллюсков.
Тела гигантских нейронов брюхоногих моллюсков — это крупные образования, не имеющие на себе синапсов. У этих нейронов синапсы находятся на так называемом центральном отростке.
Большие размеры и однородность поверхности позволяют исследовать тела этих нейронов с применением самых точных современных электрофизиологических методов. Так, с помощью двух или большего числа микроэлектродов, вводимых в эти клетки, производят электрическое раздражение клетки, регистрируют ее МПП и ПД, исследуют Rм, Сτ; производят фиксацию мембранного потенциала и на этом фоне изучают трансмембранные токи, лежащие в основе возбуждения.
В лаборатории П. Г. Костюка разработан метод внутриклеточного диализа одиночной гигантской нервной клетки. Он состоит в следующем: механически изолированную гигантскую нервную клетку помещают в соответствующем солевом растворе в камеру с плоскими стенками, на дне которой имеется коническая пора; ее широкая часть (обращенная в этот сосуд) может пропустить клетку, а узкая нет. Стенки поры покрыты клейкой массой. Под камерой, содержащей клетку, находится другая аналогичная камера (с тем же солевым раствором), имеющая сообщение с первой только через упомянутую коническую пору. Клетку подводят к поре и «засасывают» в нее, понижая гидростатическое давление в нижнем сосуде. При этом части клетки, вошедшие в пору, прилипают к ее стенкам. Часть клетки, выступающая в нижнюю камеру, разрушается пропусканием через нижнюю камеру безкальциевого раствора. После этого гидростатическое давление в нижней камере и среду в ней возвращают к норме. В итоге получается препарат соматической мембраны, омываемой снаружи жидкостью верхней камеры, а изнутри жидкостью нижней камеры. Внутриклеточные структуры при этом вымываются. На этом препарате соматической мембраны при его электрическом раздражении, как показали опыты, могут быть получены полноценные ПД. Удалось осуществить кламп потенциала такой мембраны, изучить трансмембранные токи при ее активации, а при уменьшении площади препарата даже получить характеристики токов отдельных мембранных каналов.
МПП сомы невелик (-40——50 мВ) и имеет механизм, в общем сходный с известным для аксонов. ПД имеет овершут и достигает 100 и более милливольт (мВ). По форме он схож с аксонным. Но ионные механизмы ПД мембраны сомы нейрона моллюска имеют существенные отличия от известных для нервных волокон. Главное отличие состоит в том, что ПД здесь генерируется не только за счет входящего натриевого тока (как в аксоне), но и за счет входящего кальциевого тока. В соматической мембране имеются сходные с натриевыми, но независимые от них специальные кальциевые каналы, открываемые деполяризацией. Ток Са2+ через них осуществляется по электрохимическому градиенту. Эти каналы не блокируются тетро-дотоксином (ТТХ), но блокируются кобальтом, верапамилом и препаратом Д-600. Кальциевый ток развивается медленнее натриевого и гораздо медленнее инактивируется (много секунд!).
В соматической мембране нейрона моллюсков обнаружены, кроме того, особые «быстрые» потенциалозависимые калиевые каналы (очень быстро активирующиеся и инактивирующиеся). Эти каналы почти не участвуют в формировании МПП и ПД, ибо они уже инактивированы при МП = 70 мВ, т. е. при МПП. Но эти каналы принимают участие в регулировке МП на фоне следовой гиперполяризации, тем самым определяя в какой-то мере частоту ритмического разряда в ответ на то или иное воздействие.
Калиевые каналы, участвующие в ПД, т. е. каналы «задержанного» выходящего тока, здесь также имеют особенность — они инактивируются быстрее, чем в аксоне. За счет этого у соматических ПД, следующих ритмически, прогрессивно удлиняются нисходящие фазы, но удерживается величина амплитуды, ибо ослабление калиевого шунта компенсирует ослабление натриевых токов от натриевой инактивации. Особенности ионных каналов соматической мембраны, вероятно, имеют значение не только для электрогенеза. Можно думать, что они существенны и для обеспечения интенсивно протекающих в соме метаболических реакций, чувствительных к [Са2+], [К+]и [Na+] внутри клетки. Однако у разных животных эти характеристики перикарионов не одинаковы. В телах нейронов спинномозговых ганглиев крыс, исследованных по такой же методике, не обнаружилось кальциевых каналов, участвующих в генерации ПД, вместо них там были найдены особые «медленные» натриевые каналы.
Следует сказать несколько слов о ПД перехватов Ранвье нервных волокон позвоночных животных. Потенциалы действия перехватов Ранвье имеют в общем такой же механизм, как н у гигантского аксона кальмара. Однако плотность натриевого тока ПД здесь гораздо выше. Нисходящая фаза ПД определяется в перехвате не только выходящим калиевым током, но в значительной мере неспецифическим током утечки. Роль этого тока особенно велика у птиц и млекопитающих. По последним данным у миелинизированных нервных волокон млекопитающих в мембране перехватов очень мало или даже совсем нет потенциалозависимых калиевых каналов. Но у этих волокон такие каналы имеются в межперехватной (интернодальной) мембране. По-видимому, у всех позвоночных неспецифический ток утечки, определяющий нисходящую фазу ПД в перехватах миелинизированных волокон, протекает отчасти и через насечки миелиновой оболочки. В генезе следового отрицательного потенциала неизолированного перехвата Ранвье очень большую, если не основную роль играет накопление ионов К+ в щели перехвата.
В заключение остановимся на так называемых шумах ионных каналов. Каждый из ионных каналов мембраны, обладающих теми или иными воротными механизмами, даже при постоянстве МП и химического состава среды не находится все время в каком-то одном положении; он то открывается, то закрывается. Эти переходы от закрытого состояния к открытому и обратно осуществляются по закону случая и практически мгновенно, что обусловливает прямоугольную форму одиночного толчка тока в канале. Время открытого состояния называют «временем жизни канала». Хаотическое открытие и закрытие каналов, порождающее перемещения ионов, создает электрический шум. Для потенциалозависимых каналов соответствующее электрическое поле является фактором резко увеличивающим вероятность открытого состояния каналов, что и создает эффект увеличения ионной проницаемости. Для исследования динамики отдельных каналов определенного вида исследуют слабый электрический шум, создаваемый этими каналами в каком-то небольшом участке мембраны в условиях клампа МП и химической блокады прочих каналов. Например, в мембране гигантского аксона кальмара так изучили шумы отдельно натриевых и калиевых каналов.
Анализ этих шумов позволил рассчитать плотность расположения каналов в мембране и проводимость одиночного канала.
В гигантском аксоне кальмара плотность натриевых каналов оказалась равной 300/1 мкм2, проводимость канала 4 рмО. В перехвате Ранвье амфибий плотность натриевых каналов — 2000/1 мкм2, а средняя проводимость канала равна 8 рмО.