Схема нервной системы человека
Не́рвная систе́ма
— целостная морфологическая и функциональная совокупность различных взаимосвязанных, нервных структур, которая совместно с эндокринной системой обеспечивает взаимосвязанную регуляцию деятельности всех систем организма и реакцию на изменение условий внутренней и внешней среды. Нервная система действует как интегративная система, связывая в одно целое чувствительность, двигательную активность и работу других регуляторных систем (эндокринной и иммунной).
Содержание
- 1 Общая характеристика нервной системы 1.1 Нейроны
- 1.2 Нейроглия
- 2.1 Типы нервных систем
- 2.2.6.1 Морфологическое деление
- 2.3.1 Модели 2.3.1.1 Регуляционные механизмы
- 3.1 Профессиональные сообщества и журналы
Общее развитие нервной системы
Филогенез нервной системы
* в кратких чертах сводится к следующему. У простейших одноклеточных организмов (амеба) нервной системы еще нет, а связь с окружающей средой осуществляется при помощи жидкостей, находящихся внутри и вне организма — гуморальная (humor — жидкость), донервная форма регуляции.
* (Цит. по кн.: Сепп Е. К. История развития нервной системы позвоночных от бесчерепных до человека. М., 1949.
)
В дальнейшем, когда возникает нервная система, появляется и другая форма регуляции — нервная. По мере развития нервной системы нервная регуляция все больше подчиняет себе гуморальную, так что образуется единая нейро-гуморальная регуляция при ведущей роли нервной системы. Последняя в процессе филогенеза проходит ряд основных этапов (рис. 265).
Рис. 265. Этапы развития нервной системы. 1 — диффузная нервная система; 2 — диффузная нервная система гидры; 3 и 4 — узловая нервная система кольчатого червя
I этап — сетевидная нервная система
. На этом этапе (кишечнополостные) нервная система, например гидры, состоит из нервных клеток, многочисленные отростки которых соединяются друг с другом в разных направлениях, образуя сеть, диффузно пронизывающую все тело животного. При раздражении любой точки тела возбуждение разливается по всей нервной сети, и животное реагирует движением всего тела. Отражением этого этапа у человека является сетевидное строение интрамуральной нервной системы.
II этап — узловая нервная система
. На этом этапе (высшие черви) нервные клетки сближаются в отдельные скопления или группы, причем из скоплений клеточных тел получаются нервные узлы — центры, а из скоплений отростков — нервные стволы — нервы. При этом в каждой клетке число отростков уменьшается, и они получают определенное направление. Соответственно сегментарному строению тела животного, например у кольчатого червя, в каждом сегменте имеются сегментарные нервные узлы и нервные стволы. Последние соединяют узлы в двух направлениях поперечные стволы связывают узлы данного сегмента, а продольные — узлы разных сегментов. Благодаря этому нервные импульсы, возникающие в какой-либо точке тела, не разливаются по всему телу, а распространяются по поперечным стволам в пределах данного сегмента. Продольные стволы связывают нервные сегменты в одно целое. На головном конце животного, который при движении вперед соприкасается с различными предметами окружающего мира, развиваются органы чувств, в связи с чем головные узлы развиваются сильнее остальных, являясь прообразом будущего головного мозга. Отражением этого этапа является сохранение у человека примитивных черт (разбросанность на периферии узлов и микроганглиев) в строении вегетативной нервной системы.
III этап — трубчатая нервная система
. На первоначальной ступени развития животных особенно большую роль играл аппарат движения, от совершенства которого зависит основное условие существования животного — питание (передвижение в поисках пищи, захватывание и поглощение ее).
У низших многоклеточных развился перистальтический способ передвижения, что связано с гладкой мускулатурой и ее местным нервным аппаратом. На более высокой ступени перистальтический способ сменяется скелетной моторикой, т. е. передвижением с помощью системы жестких рычагов- поверх мышц (членистоногие) и внутри мышц (позвоночные). Следствием этого явилось образование поперечнополосатой мускулатуры и центральной нервной системы, координирующей перемещение отдельных рычагов моторного скелета.
Такая центральная нервная система у хордовых (ланцетник) возникла в виде метамерно построенной нервной трубки с отходящими от нее сегментарными нервами ко всем сегментам тела, включая и аппарат движения — туловищный мозг. У позвоночных и человека туловищный мозг становится спинным. Таким образом, появление туловищного мозга связано с усовершенствованием в первую очередь моторного вооружения животного. Наряду с этим уже у ланцетника имеются и рецепторы (обонятельный, световой). Дальнейшее развитие нервной системы и возникновение головного мозга обусловлены преимущественно усовершенствованием рецепторного вооружения.
Так как большинство органов чувств возникает на том конце тела животного, который обращен в сторону движения, т. е. вперед, то для восприятия поступающих через них внешних раздражений развивается передний конец туловищного мозга и образуется головной мозг, что совпадает с обособлением переднего конца тела в виде головы — цефализация (cephal — голова).
Е. К. Сепп в руководстве по нервным болезням* дает упрощенную, но удобную для изучения схему филогенеза головного мозга, которую мы и приводим. Согласно этой схеме, на первом этапе развития головной мозг состоит из трех отделов: заднего, среднего и переднего, причем из этих отделов в первую очередь (у низших рыб) особенно развивается задний, или ромбовидный, мозг (rhombencephalon). Развитие заднего мозга происходит под влиянием рецепторов акустики и статики (рецепторы VIII пары головных нервов), имеющих ведущее значение для ориентации в водной среде.
* (Е. К. Сепп, М. Б. Цукер, Е. В. Шмидт, 1950.
)
В дальнейшей эволюции задний мозг дифференцируется на продолговатый мозг
, являющийся переходным отделом от спинного мозга к головному и потому называемый myelencephalon (myelos — спинной мозг, encephalon — головной), и
собственно задний мозг
— metencephalon, из которого развиваются мозжечок и мост.
В процессе приспособления организма к окружающей среде путем изменения обмена веществ в заднем мозгу как наиболее развитом на этом этапе отделе центральной нервной системы возникают центры управления жизненно важными процессами растительной жизни, связанными, в частности, с жаберным аппаратом (дыхание, кровообращение, пищеварение и др.). Поэтому в продолговатом мозгу возникают ядра жаберных нервов (группа X пары — вагуса). Эти жизненно важные центры дыхания и кровообращения остаются в продолговатом мозгу человека, чем объясняется смерть, наступающая при повреждении продолговатого мозга. На II этапе (еще у рыб) под влиянием зрительного рецептора особенно развивается средний мозг
, mesencephalon. На III этапе, в связи с окончательным переходом животных из водной среды в воздушную, усиленно развивается обонятельный рецептор, воспринимающий содержащиеся в воздухе химические вещества, сигнализирующие своим запахом о добыче, опасности и других жизненно важных явлениях окружающей природы.
Под влиянием обонятельного рецептора развивается передний мозг — prosencephalon, вначале имеющий характер чисто обонятельного мозга. В дальнейшем передний мозг разрастается и дифференцируется на промежуточный
— diencephalon и
конечный
— telencephalon.
В конечном мозгу как в высшем отделе центральной нервной системы появляются центры для всех видов чувствительности. Однако нижележащие центры не исчезают, а сохраняются, подчиняясь центрам вышележащего этажа. Следовательно, с каждым новым этапом развития головного мозга возникают новые центры, подчиняющие себе старые. Происходит как бы передвижение функциональных центров к головному концу и одновременное подчинение филогенетически старых зачатков новым. В результате центры слуха, впервые возникшие в заднем мозгу, имеются также в среднем и переднем, центры зрения, возникшие в среднем, имеются и в переднем, а центры обоняния — только в переднем мозгу. Под влиянием обонятельного рецептора развивается небольшая часть переднего мозга, называемая поэтому обонятельным мозгом (rhinencephalon), который покрыт корой серого вещества — старой корой (paleocortex).
Совершенствование рецепторов приводит к прогрессивному развитию переднего мозга, который постепенно становится органом, управляющим всем поведением животного. Различают две формы поведения животного: инстинктивное, основанное на видовых реакциях (безусловные рефлексы), и индивидуальное, основанное на опыте индивида (условные рефлексы). Соответственно этим двум формам поведения в конечном мозгу развиваются две группы центров серого вещества: подкорковые узлы, имеющие строение ядер (ядерные центры), и кора серого вещества, имеющая строение сплошного экрана (экранные центры). При этом вначале развивается «подкорка», а затем кора. Кора возникает при переходе животного от водного к наземному образу жизни и обнаруживается отчетливо у амфибий и рептилий. Дальнейшая эволюция нервной системы характеризуется тем, что кора головного мозга все более и более подчиняет себе функции всех нижележащих центров, происходит постепенная кортиколизация функций
.
Необходимой формацией для осуществления высшей нервной деятельности является новая кора, расположенная на поверхности полушарий и приобретающая в процессе филогенеза шестислойное строение. Благодаря усиленному развитию новой коры конечный мозг у высших позвоночных превосходит все остальные отделы головного мозга, покрывая их, как плащом (pallium). Развивающийся новый мозг (neencephalon) оттесняет в глубину старый мозг (обонятельный), который как бы свертывается в виде аммонова рога (cornu Ammoni или pes hyppocampi), остающегося по-прежнему обонятельным центром. В результате плащ, т. е. новый мозг
(neencephalon), резко преобладает над остальными отделами мозга — старым мозгом (paleencephalon).
Итак, развитие головного мозга совершается под влиянием развития рецепторов, чем и объясняется, что самый высший отдел головного мозга — кора серого вещества — представляет, как учит И. П. Павлов, совокупность корковых концов анализаторов, т. е. сплошную воспринимающую (рецепторную) поверхность. Дальнейшее развитие мозга у человека подчиняется иным закономерностям, связанным с его социальной природой. Кроме естественных органов тела, имеющихся и у животных, человек стал пользоваться орудиями труда. Орудия труда, ставшие искусственными органами, дополнили естественные органы тела и составили техническое вооружение человека.
С помощью этого вооружения человек приобрел возможность не только приспосабливаться самому к природе, как это делают животные, но и приспосабливать природу к своим нуждам. Труд, как уже говорилось выше, явился решающим фактором становления человека, а в процессе общественного труда возникло необходимое для общения людей средство — речь. «Сначала труд, а затем и вместе с ним членораздельная речь явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны постепенно превратился в человеческий мозг, который, при всем своем сходстве с обезьяньим, далеко превосходит его по величине и совершенству» (К. Маркс, Ф. Энгельс. Сочинения, изд. 2, т. 20, с. 490). Это совершенство обусловлено максимальным развитием конечного мозга, особенно его коры — новой коры (neocortex).
Кроме анализаторов, воспринимающих различные раздражения внешнего мира и составляющих материальный субстрат конкретно-наглядного мышления, свойственного животным (первая сигнальная система
действительности, по И. П. Павлову), у человека возникла способность абстрактного, отвлеченного мышления с помощью слова, сначала слышимого (устная речь) и позднее видимого (письменная речь). Это составило
вторую сигнальную систему
, по И. П. Павлову, которая в развивающемся животном мире явилась «чрезвычайной прибавкой к механизмам нервной деятельности» (И. П. Павлов). Материальным субстратом второй сигнальной системы стали поверхностные слои новой коры. Поэтому кора конечного мозга достигает своего наивысшего развития у человека. Таким образом, вся эволюция нервной системы сводится к прогрессивному развитию конечного мозга, который у высших позвоночных и особенно у человека в связи с усложнением нервных функций достигает огромных размеров.
Изложенные закономерности филогенеза обусловливают эмбриогенез нервной системы
человека. Нервная система происходит из наружного зародышевого листка, или эктодермы (см. «Введение»). Эта последняя образует продольное утолщение, называемое
медуллярной пластинкой
(рис. 266). Медуллярная пластинка скоро углубляется в
медуллярную бороздку
, края которой (медуллярные валики) постепенно становятся выше и затем срастаются друг с другом, превращая бороздку в трубку (
мозговая трубка
). Мозговая трубка представляет собой зачаток центральной части нервной системы. Задний конец трубки образует зачаток спинного мозга, передний же ее расширенный конец путем перетяжек расчленяется на три первичных мозговых пузырька, из которых происходит головной мозг во всей его сложности.
Рис. 266. Стадии эмбриогенеза нервной системы в поперечном схематическом разрезе. А — медуллярная пластинка; В и С — медуллярная бороздка; D и Е — мозговая трубка; 1 — роговой листок (эпидермис); 2 — ганглиозный валик
Мозговая пластинка первоначально состоит только из одного слоя эпителиальных клеток. Во время замыкания ее в мозговую трубку количество клеток в стенках последней увеличивается, так что возникает три слоя: внутренний (обращенный в полость трубки), из которого происходит эпителиальная выстилка мозговых полостей (эпендима центрального канала спинного мозга и желудочков головного); средний, из которого развивается серое вещество мозга (нервные клетки — нейробласты), наконец, наружный, почти не содержащий клеточных ядер, развивающийся в белое вещество (отростки нервных клеток — нейриты). Пучки нейритов нейробластов распространяются или в толще мозговой трубки, образуя белое вещество мозга, или же выходят в мезодерму и затем соединяются с молодыми мышечными клетками (миобластами). Таким путем возникают двигательные нервы.
Чувствительные нервы возникают из зачатков спинномозговых узлов, которые заметны уже по краям медуллярной бороздки у места перехода ее в кожную эктодерму. Когда бороздка смыкается в мозговую трубку, зачатки смещаются на ее дорсальную сторону, располагаясь по средней линии. Затем клетки этих зачатков перемещаются вентрально и располагаются вновь по бокам мозговой трубки в виде так называемых ганглиозных валиков
. Оба ганглиозных валика перешнуровываются четкообразно по сегментам дорсальной стороны зародыша, вследствие чего получается на каждой стороне ряд спинномозговых узлов, ganglia spinalia s. intervertebralia. В головной части мозговой трубки они доходят только до области заднего мозгового пузырька, где образуют зачатки узлов чувствительных головных нервов. В ганглиозных зачатках развиваются нейробласты, принимающие вид биполярных нервных клеток, один из отростков которых врастает в мозговую трубку, другой идет на периферию, образуя чувствительный нерв. Благодаря сращению на некотором протяжении от начала обоих отростков получаются из биполярных так называемые ложные униполярные клетки с одним отростком, делящимся в форме буквы «Т», являющиеся характерными для межпозвонковых узлов взрослого. Центральные отростки клеток, проникающие в спинной мозг, составляют задние корешки спинномозговых нервов, а периферические отростки, разрастаясь вентрально, образуют (вместе с вышедшими из спинного мозга эфферентными волокнами, составляющими передний корешок) смешанный спинномозговой нерв. Из ганглиозных валиков возникают также зачатки вегетативной нервной системы.
Общая характеристика нервной системы
Всё разнообразие значений нервной системы вытекает из её свойств.
- Возбудимость, раздражимость и проводимость характеризуются как функции времени, то есть это — процесс, возникающий от раздражения до проявления ответной деятельности органа. Согласно электрической теории распространения нервного импульса в нервном волокне, он распространяется за счёт перехода локальных очагов возбуждения на соседние неактивные области нервного волокна или процесса распространяющейся деполяризации потенциала действия, представляющего подобие электрического тока. В синапсах протекает другой — химический процесс, при котором развитие волны возбуждения-поляризации принадлежит медиатору ацетилхолину, то есть химической реакции.
- Нервная система обладает свойством трансформации и генерации энергий внешней и внутренней среды и преобразования их в нервный процесс.
- К особенно важному свойству нервной системы относится свойство мозга хранить информацию в процессе не только онто-, но и филогенеза.
Декарт: «Раздражение ступни передаётся по нервам в мозг, взаимодействует там с духом и таким образом порождает ощущение боли».
Нейроны
Основная статья: Нейрон
Нервная система состоит из нейронов, или нервных клеток и нейроглии, или нейроглиальных (или глиальных) клеток. Нейроны
— это основные структурные и функциональные элементы как в центральной, так и периферической нервной системе. Нейроны — это возбудимые клетки, то есть они способны генерировать и передавать электрические импульсы (потенциалы действия). Нейроны имеют различную форму и размеры, формируют отростки двух типов:
аксоны
и
дендриты
. Дендритов может быть много, несколько, один или не быть вообще. Обычно у нейрона несколько коротких разветвлённых дендритов, по которым импульсы следуют к телу нейрона, и всегда один длинный аксон, по которому импульсы идут от тела нейрона к другим клеткам (нейронам, мышечным либо железистым клеткам). Нейроны, по форме и характеру отхождения от них отростков, бывают: униполярные (одноотростковые), биополярные (двуотростковые), псевдоуниполярные (ложноотростковые) и мультиполярные (многоотростковые). По размерам нейроны бывают: мелкие (до 5 мкм), средние (до 30 мкм) и крупные (до 100 мкм). Длина отростков у нейронов различна: например, у одних длина отростков микроскопическая, а у других до 1,5 м. Так, например, нейрон находится в спинном мозге, а его отростки заканчиваются в пальцах рук или ног. Передача нервного импульса (возбуждения), а также регуляция его интенсивности, с одного нейрона на другие клетки происходит посредством специализированных контактов — синапсов.
Нейроглия
Основная статья: Нейроглия
Глиальные клетки
более многочисленны, чем нейроны и составляют по крайней мере половину объёма ЦНС, но в отличие от нейронов они не могут генерировать потенциалов действия. Нейроглиальные клетки различны по строению и происхождению, они выполняют вспомогательные функции в нервной системе, обеспечивая опорную, трофическую, секреторную, разграничительную и защитную функции.
Строение и функции зрительного нерва
Сам по себе зрительный нерв — это сплетение множества тончайших нервных волокон, между которыми находится центральный артериальный канал сетчатки. Нервные волокна собираются в зрительный нерв у заднего полюса глаза. В месте, где они приближаются к диску зрительного нерва (ДЗН), количество волокон увеличивается, поэтому здесь образуется небольшое возвышение над сетчаткой. Далее волокна собираются в диск, изгибаются под углом 90˚ и ограничивают внутриглазной отдел зрительного нерва.
У зрительного нерва три оболочки:
- Твердая (мозговая) — соединительнотканная пластинка, толщиной не более 0,35-0,50 мм, которая значительно утолщается в месте ее перехода в склеру
- Паутинная — тонкий слой (толщина всего 10 мкм) коллагеновой ткани, покрытой плоскими клетками.
- Мягкая — рыхлая соединительная ткань, в состав которой входят эластичные, коллагеновые, ретикулярные волокна, а также фибробласты.
Пространство между ними заполняется жидкостью со сложным составом.
Зрительный нерв делят на четыре участка: внутриглазной, внутриорбитальный, внутриканальцевый и внутричерепной.
Начинается зрительный нерв в диске зрительного нерва, а заканчивается в хиазме — своеобразном перекресте нервов. После этого часть нервных волокон проходит в соответствующие центры мозга.
Так зрительный нерв выполняет свою основную функцию — доставку первичных импульсов к отсекам головного мозга. Отсюда импульсы возвращают в зрительный центр уже готовое изображение.
Сравнительная нейроанатомия
Типы нервных систем
Существует несколько типов организации нервной системы, представленные у различных систематических групп животных.
- Диффузная нервная система — представлена у кишечнополостных. Нервные клетки образуют диффузное нервное сплетение в эктодерме по всему телу животного, и при сильном раздражении одной части сплетения возникает генерализованный ответ — реагирует все тело.
- Стволовая нервная система (ортогон) — некоторые нервные клетки собираются в нервные стволы, наряду с которыми сохраняется и диффузное подкожное сплетение. Такой тип нервной системы представлен у плоских червей и нематод (у последних диффузное сплетение сильно редуцировано), а также многих других групп первичноротых — например, гастротрих и головохоботных.
- Узловая нервная система, или сложная ганглионарная система — представлена у аннелид, членистоногих, моллюсков и других групп беспозвоночных. Большая часть клеток центральной нервной системы собраны в нервные узлы — ганглии. У многих животных клетки в них специализированы и обслуживают отдельные органы. У некоторых моллюсков (например, головоногих) и членистоногих возникает сложное объединение специализированных ганглиев с развитыми связями между ними — единый головной мозг или головогрудная нервная масса (у пауков). У насекомых особенно сложное строение имеют некоторые отделы протоцеребрума («грибовидные тела»).
- Трубчатая нервная система (нервная трубка) характерна для хордовых.
Нервная система различных животных
Нервная система книдарий и гребневиков
Наиболее примитивными животными, у которых есть нервная система, считаются книдарии. У полипов она представляет собой примитивную субэпителиальную нервную сеть (нервный плексус
), оплетающую всё тело животного и состоящую из нейронов разного типа (чувствительных и ганглиозных клеток), соединённых друг с другом отростками (
диффузная нервная система
), особенно плотные их сплетения образуются на оральном и аборальном полюсах тела. Раздражение вызывает быстрое проведение возбуждения по телу гидры и приводит к сокращению всего тела, в связи с сокращением эпителиально-мускульных клеток эктодермы и одновременно их расслаблением в энтодерме. Медузы устроены сложнее полипов, в их нервной системе начинает обособляться центральный отдел. Помимо подкожного нервного сплетения у них имеются ганглии по краю зонтика, соединённые отростками нервных клеток в
нервное кольцо
, от которого иннервируются мышечные волокна паруса и
ропалии
— структуры, содержащие различные органы чувств (
диффузно-узловая нервная система
). Бо́льшая централизация наблюдается у сцифомедуз и особенно кубомедуз. Их 8 ганглиев, соответствующие 8 ропалиям, достигают достаточно крупных размеров.
Нервная система гребневиков включает субэпителиальное нервное сплетение со сгущениями вдоль рядов гребных пластинок, которые сходятся к основанию сложно устроенного аборального органа чувств. У некоторых гребневиков описаны находящиеся рядом с ним нервные ганглии.
Нервная система первичноротых
Плоские черви
имеют уже подразделённую на центральный и периферический отделы нервную систему. В целом нервная система напоминает правильную решётку — такой тип строения был назван
ортогоном
. Она состоит из мозгового ганглия, у многих групп окружающего статоцист (эндонного мозга), который соединен с
нервными стволами
ортогона, идущими вдоль тела и соединённые кольцевыми поперечными перемычками (
комиссурами
). Нервные стволы состоят из нервных волокон, отходящих от рассеянных по их ходу нервных клеток. У некоторых групп нервная система довольно примитивна и близка к диффузной. Среди плоских червей наблюдаются следующие тенденции: упорядочивание подкожного сплетения с обособлением стволов и комиссур, увеличение размеров мозгового ганглия, который превращается в центральный аппарат управления, погружение нервной системы в толщу тела; и, наконец, уменьшение числа нервных стволов (у некоторых групп сохраняются лишь два
брюшных (боковых) ствола
).
У немертин центральная часть нервной системы представлена парой соединённых двойных ганглиев, расположенных над и под влагалищем хоботка, соединённых комиссурами и достигающих значительного размера. От ганглиев идут назад нервные стволы, обычно их пара и расположены они по бокам тела. Они также соединены комиссурами, расположены они в кожно-мускульном мешке или в паренхиме. От головного узла отходят многочисленные нервы, наиболее сильно развиты спинной нерв (часто двойной), брюшной и глоточный.
У брюхоресничных червей имеется надглоточный ганглий, окологлоточное нервное кольцо и два поверхностных боковых продольных ствола, соединённых комиссурами.
У нематод имеется окологлоточное нервное кольцо, вперёд и назад от которого отходят по 6 нервных стволов, наиболее крупные — брюшной и спинной стволы — тянутся вдоль соответствующих гиподермальных валиков. Между собой нервные стволы связаны полукольцевыми перемычками, иннервируют они соответственно мышцы брюшных и спинных боковых лент. Нервная система нематоды Caenorhabditis elegans
была закартирована на клеточном уровне[1]. Каждый нейрон был зарегистрирован, прослежено его происхождение и большинство, если не все, нейронные связи известны[2]. У этого вида нервная система обладает половым диморфизмом: мужская и гермафродитная нервная система имеют разное количество нейронов и групп нейронов, чтобы выполнять полоспецифические функции.
У киноринх нервная система состоит из окологлоточного нервного кольца и вентрального (брюшного) ствола, на котором, в соответствии с присущей им сегментацией тела, группами расположены ганглионарные клетки.
Схоже устроена нервная система волосатиков и приапулид, но их вентральный нервный ствол лишен утолщений.
У коловраток имеется крупный надглоточный ганглий, от которого отходят нервы, особенно крупные — два нерва, идущие через всё тело по бокам кишечника. Более мелкие ганглии лежат в ноге (педальный ганглий) и рядом с жевательным желудком (ганглий мастакса).
У скребней нервная система очень проста: внутри влагалища хоботка имеется непарный ганглий, от которого отходят тонкие веточки вперёд к хоботку и два более толстых боковых ствола назад, они выходят из влагалища хоботка, пересекают полость тела, а затем по её стенкам идут назад.
У кольчатых червей имеется парный надглоточный нервный узел, окологлоточными коннективами
(коннективы в отличие от комиссур соединяют разноимённые ганглии) соединённый с брюшной частью нервной системы. У примитивных полихет она состоит из двух продольных нервных тяжей, в которых располагаются нервные клетки. У более высокоорганизованных форм они образуют парные ганглии в каждом сегменте тела (
нервная лестница
), а нервные стволы сближаются. У большинства же полихет парные ганглии сливаются (
брюшная нервная цепочка
), у части сливаются и их коннективы. От ганглиев отходят многочисленные нервы к органам своего сегмента. В ряду полихет происходит погружение нервной системы из-под эпителия в толщу мышц или даже под кожно-мускульный мешок. Ганглии разных сегментов могут концентрироваться, если сливаются их сегменты. Аналогичные тенденции наблюдаются и у олигохет. У пиявок нервная цепочка, лежащая в брюшном лакунарном канале, состоит из 20 или более ганглиев, причём в один объединяются первые 4 ганглия (
подглоточный нервный узел
) и последние 7.
У эхиурид нервная система развита слабо — окологлоточное нервное кольцо соединено с брюшным стволом, но нервные клетки рассеяны по ним равномерно и нигде не образуют узлов.
У сипункулид имеется надглоточный нервный ганглий, окологлоточное нервное кольцо и лишённый нервных узлов брюшной ствол, лежащий на внутренней стороне полости тела.
Тихоходки имеют надглоточный ганглий, окологлоточные коннективы и брюшную цепочку с 5 парными ганглиями.
Онихофоры имеют примитивную нервную систему. Мозг состоит из трёх отделов: протоцеребрум иннервирует глаза, дейтоцеребрум — антенны, а тритоцеребрум — переднюю кишку. От окологлоточных коннектив отходят нервы к челюстям и ротовым сосочкам, а сами коннективы переходят в далёкие друг от друга брюшные стволы, равномерно покрытые нервными клетками и соединённые тонкими комиссурами.
Нервная система членистоногих
У членистоногих нервная система слагается из парного надглоточного узла, состоящего из нескольких соединённых нервных узлов (головной мозг), окологлоточных коннектив и брюшной нервной цепочки, состоящей из двух параллельных стволов. У большинства групп головной мозг делится на три отдела — прото-
,
дейто-
и
тритоцеребрум
. Каждый сегмент тела имеет по паре нервных ганглиев, но часто наблюдается слияние ганглиев с образованием крупных нервных центров; например, подглоточный нервный узел состоит из нескольких пар сросшихся ганглиев — он контролирует слюнные железы и некоторые мышцы пищевода.
В ряду ракообразных в целом наблюдаются те же тенденции, что и у кольчатых червей: сближение пары брюшных нервных стволов, слияние парных узлов одного сегмента тела (то есть образование брюшной нервной цепочки), слияние её узлов в продольном направлении по мере объединения сегментов тела. Так, у крабов имеется лишь две нервные массы — головной мозг и нервная масса в груди, а у веслоногих и ракушковых раков образуется единственное компактное образование, пронизанное каналом пищеварительной системы. Головной мозг раков состоит из парных долей — протоцеребрума, от которого отходят зрительные нервы, имеющие ганглиозные скопления нервных клеток, и дейтоцеребрума, иннервирующего антенны I. Обычно добавляется и тритоцеребрум, образованный слившимися узлами сегмента антенн II, нервы к которым обычно отходят от окологлоточных коннективов. У ракообразных имеется развитая симпатическая нервная система
, состоящая из мозгового отдела и непарного
симпатического нерва
, имеющего несколько ганглиев и иннервирующего кишечник. Важную роль в физиологии раков играют
нейросекреторные клетки
, расположенные в различных частях нервной системы и выделяющие
нейрогормоны
.
Головной мозг многоножек имеет сложное строение, образован, скорее всего, многими ганглиями. Подглоточный ганглий иннервирует все ротовые конечности, от него начинается длинный парный продольный нервный ствол, на котором в каждом сегменте приходится по одному парному ганглию (у двупарноногих многоножек в каждом сегменте, начиная с пятого, по две пары ганглиев, расположенных одна за другой).
Нервная система насекомых, также состоящая из головного мозга и брюшной нервной цепочки, может достигать значительного развития и специализации отдельных элементов. Головной мозг состоит из трёх типичных отделов, каждый из которых состоит из нескольких ганглиев, разделённых прослойками нервных волокон. Важным ассоциативным
протоцеребрума. Особенно развитый мозг у общественных насекомых (муравьёв, пчёл, термитов). Брюшная нервная цепочка состоит из подглоточного нервного узла, иннервирующего ротовые конечности, трёх крупных грудных узлов и брюшных узлов (не более 11). У большинства видов не встречается во взрослом состоянии более 8 ганглиев, у многих и они сливаются, давая крупные ганглиозные массы. Может доходить до образования только одной ганглиозной массы в груди, иннервирующей и грудь, и брюшко насекомого (например, у некоторых мух). В онтогенезе зачастую происходит объединение ганглиев. От головного мозга отходят симпатические нервы. Практически во всех отделах нервной системы имеются нейросекреторные клетки.
У мечехвостов головной мозг внешне не расчленён, но имеет сложное гистологическое строение. Утолщённые окологлоточные коннективы иннервируют хелицеры, все конечности головогруди и жаберные крышки. Брюшная нервная цепочка состоит из 6 ганглиев, задний образован слиянием нескольких. Нервы брюшных конечностей соединены продольными боковыми стволами.
Нервная система паукообразных имеет чёткую тенденцию к концентрации. Головной мозг состоит только из протоцеребрума и тритоцеребрума в связи с отсутствием структур, которые иннервирует дейтоцеребрум. Метамерность брюшной нервной цепочки яснее всего сохраняется у скорпионов — у них большая ганглиозная масса в груди и 7 ганглиев в брюшке, у сольпуг их только 1, а у пауков все ганглии слились в головогрудную нервную массу; у сенокосцев и клещей нет разграничения между нею и головным мозгом.
Морские пауки, как и все хелицеровые, не имеют дейтоцеребрума. Брюшная нервная цепочка у разных видов содержит от 4—5 ганглиев до одной сплошной ганглиозной массы.
Нервная система моллюсков
У примитивных моллюсков хитонов нервная система состоит из окологлоточного кольца (иннервирует голову) и 4 продольных стволов — двух педальных
(иннервируют ногу, которые связаны без особого порядка многочисленными комиссурами, и двух
плевровисцеральных
, которые расположены кнаружи и выше педальных (иннервируют внутренностный мешок, над порошицей соединяются). Педальный и плевровисцеральный стволы одной стороны также связаны множеством перемычек.
Схоже устроена нервная система моноплакофор, но педальные стволы соединяются у них только одной перемычкой.
У более развитых форм образуется в результате концентрации нервных клеток несколько пар ганглиев, которые смещаются к переднему концу тела, причём наибольшее развитие получает надглоточный узел (головной мозг).
Нервная система вторичноротых
Нервная система позвоночных
Нервная система позвоночных животных часто делится на центральную нервную систему (ЦНС) и периферическую нервную систему (ПНС) . ЦНС состоит из головного и спинного мозга. ПНС состоит из других нервов и нейронов, которые не лежат в пределах ЦНС. Преобладающее большинство нервов (которые фактически являются аксонами нейронов) принадлежит ПНС. Периферическая нервная система делится на соматическую нервную систему и вегетативную нервную систему.
Соматическая нервная система отвечает за координацию движения тела, а также за получение и передачу внешних стимулов . Эта система регулирует действия, которые находятся под сознательным контролем.
Автономная нервная система делится на парасимпатический и симпатический. Симпатическая нервная система отвечает опасности или стресс, и, среди многих физиологических изменений, может вызвать увеличение сердечного ритма и кровяного давления и возбуждения органов чувств благодаря увеличению адреналина в крови. Парасимпатическая нервная система, с другой стороны, отвечает за состояние отдыха, и обеспечивает сокращение зрачка, замедление сердца, расширение кровеносных сосудов и стимулирования пищеварительных и мочеполовых систем .
Нервная система млекопитающих
Нервная система функционирует как единое целое с органами чувств, например глазами, и управляется у млекопитающих головным мозгом. Самая крупная часть последнего называется большими полушариями (в затылочной области черепа находятся два более мелких полушария мозжечка) . Головной мозг соединяется со спинным. У всех млекопитающих, за исключением однопроходных и сумчатых, в отличие от других позвоночных правое и левое большие полушария соединены между собой компактным пучком нервных волокон, называемым мозолистым телом. В мозгу однопроходных и сумчатых мозолистого тела нет, но соответствующие области полушарий тоже соединяются нервными пучками; например, передняя комиссура связывает друг с другом правую и левую обонятельные области. Спинной мозг — основной нервный ствол тела — проходит через канал, образованный отверстиями позвонков, и тянется от головного мозга до поясничного или крестцового отдела позвоночника, в зависимости от вида животного. С каждой стороны спинного мозга симметрично отходят нервы к различным частям тела. Осязание в общих чертах обеспечивается определёнными нервными волокнами, бесчисленные окончания которых находятся в коже. Эта система обычно дополняется волосками, действующими, как рычаги, для нажатия на пронизанные нервами участки.
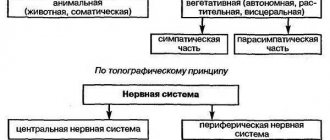
Морфологическое деление
Нервная система млекопитающих и человека по морфологическим признакам подразделяется на центральную (головной и спинной мозг) и периферическую (слагается отходящими от головного и спинного мозга нервами[3]).
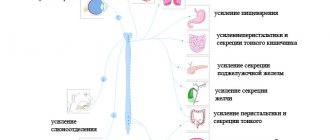
Состав центральной нервной системы можно представить следующим образом:
| |||||||||||||||||||
К периферической нервной системе относят черепные нервы, спинномозговые нервы и нервные сплетения
Функциональное деление
- Соматическая (анимальная) нервная система
- Автономная (вегетативная) нервная система Симпатический отдел вегетативной нервной системы
- Парасимпатический отдел вегетативной нервной системы
- Метасимпатический отдел вегетативной нервной системы (энтеральная нервная система)
Онтогенез
Модели
В настоящий момент нет единого положения о развитии нервной системы в онтогенезе. Основная проблема заключается в оценке уровня детерминированности (предопределения) в развитии тканей из зародышевых клеток. Наиболее перспективными моделями являются мозаичная модель
и
регуляционная модель
. Ни та, ни другая не может в полной мере объяснить развитие нервной системы.
- Мозаичная модель предполагает полное детерминирование судьбы отдельной клетки на протяжении всего онтогенеза.
- Регуляционная модель предполагает случайное и изменяемое развитие отдельных клеток, при детерминированности только нейрального направления (то есть любая клетка определённой группы клеток может стать какой угодно в пределах возможности развития для этой группы клеток).
Для беспозвоночных мозаичная модель практически безупречна — степень детерминации их бластомеров очень высока. Но для позвоночных все гораздо сложнее. Некая роль детерминации и здесь несомненна. Уже на шестнадцатиклеточной стадии развития бластулы позвоночных можно с достаточной долей уверенности сказать, какой бластомер не является
предшественником определённого органа.
Маркус Джакобсон в 1985 году ввел клональную модель развития головного мозга (близка к регуляционной). Он предположил, что детерминирована судьба отдельных групп клеток, представляющих собой потомство отдельного бластомера, то есть, «клонов» этого бластомера. Муди и Такасаки (независимо) развили эту модель в 1987. Построена карта 32-клеточной стадии развития бластулы. Например, установлено, что потомки бластомера D2 (вегетативный полюс) всегда встречаются в продолговатом мозге. С другой стороны, потомки почти всех бластомеров анимального полюса не имеют выраженной детерминации. У разных организмов одного вида они могут встречаться или не встречаться в определённых отделах головного мозга.
Регуляционные механизмы
Выяснено, что развитие каждого бластомера зависит от наличия и концентрации специфических веществ — паракринных факторов, которые выделяются другими бластомерами. Например в опыте in vitro
с апикальной частью бластулы оказалось, что в отсутствие активина (паракринного фактора вегетативного полюса) клетки развиваются в обычный эпидермис, а при его наличии, в зависимости от концентрации, по возрастанию её: клетки мезенхимы, гладкомышечные, клетки хорды или клетки сердечной мышцы.
Все вещества, определяющие поведение и судьбу клеток, их воспринимающих, в зависимости от дозы (концентрации) вещества в данном участке многоклеточного зародыша, называются морфогенами
.
Одни клетки секретируют во внеклеточное пространство растворимые активные молекулы (морфогены), убывающие от своего источника по градиенту концентрации.
Та группа клеток, чьё расположение и назначение задано в пределах одних и тех же границ (с помощью морфогенов), называется морфогенетическим полем
. Судьба самого морфогенетического поля жестко определена. Каждое конкретное морфогенетическое поле отвечает за образование конкретного органа, даже если эту группу клеток трансплантировать в различные части зародыша. Судьбы же отдельных клеток внутри поля зафиксированы не столь жестко, так что они могут в известных пределах менять назначение, восполняя функции утраченных полем клеток. Концепция морфогенетического поля является более общим понятием, по отношению к нервной системе она отвечает регуляторной модели.
С понятиями морфоген и морфогенетическое поле тесно связано понятие эмбриональной индукции. Это явление, также общее для всех систем организма, впервые было показано на развитии нервной трубки.
Развитие нервной системы позвоночных
Нервная система образуется из эктодермы — наружного из трёх зародышевых листков. Между клетками мезодермы и эктодермы начинается паракринное взаимодействие, то есть в мезодерме вырабатывается специальное вещество — фактор роста нейронов, которое передаётся в эктодерму. Под влиянием фактора роста нейронов часть эктодермальных клеток превращается в нейроэпиталиальные клетки, причём образование нейроэпителиальных клеток происходит очень быстро — со скоростью 250000 штук в минуту. Этот процесс называется нейрональной индукцией (частный случай эмбриональной индукции).
В результате образуется нервная пластинка, которая состоит из одинаковых клеток. Из неё образуются нервные валики, а из них — нервная трубка, которая обособляется от эктодермы (конкретно за образование нервной трубки и нервного гребня отвечает смена типов кадгеринов, молекул клеточной адгезии), уходя под неё. Механизм нейруляции несколько различается у низших и высших позвоночных. Замыкается нервная трубка не одновременно по всей длине. Прежде всего замыкание происходит в средней части, затем этот процесс распространяется к заднему и переднему её концам. На концах трубки сохраняется два незамкнутых участка — передний и задний нейропоры.
Затем происходит процесс дифференциации нейроэпителиальных клеток на нейробласты и глиобласты. Глиобласты дают начало астроцитам, олигодендроцитам и эпиндимным клеткам. Нейробласты становятся нейронами. Далее происходит процесс миграции — нейроны перемещаются туда, где они будут выполнять свою функцию. За счёт конуса роста нейрон ползет, подобно амёбе, а путь ему указывают отростки глиальных клеток. Следующий этап — агрегация (слипание однотипных нейронов, например, участвующих в образовании мозжечка, таламуса и пр). Нейроны узнают друг друга благодаря поверхностным лигандам — специальным молекулам, имеющимся на их мембранах. Объединившись, нейроны выстраиваются в необходимом для данной структуры порядке.
После этого идёт созревание нервной системы. Из конуса роста нейрона вырастает аксон, от тела отрастают дендриты.
Затем происходит фасцикуляция — объединение однотипных аксонов (образование нервов).
Последний этап — запрограммированная гибель тех нервных клеток, в которых произошёл сбой за время формирования нервной системы (около 8 % клеток посылают свой аксон не туда, куда нужно).
3.1. Нервная система беспозвоночных животных
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных.
Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют
плазматические контакты
, обеспечивающие непрерывность сети (
анастомозы
). Появляются и
щелевидные контакты
между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые
симметричные синапсы
, а есть и
несимметричные синапсы:
в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей.
Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис.
9
,
3
,
4
).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4
,
5
) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10,
6
). Упорядоченная таким образом нервная система называется
ортогоном.
Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
Наряду с двухсторонней симметрией у плоских червей оформляется передний конец тела, на котором концентрируются органы чувств (статоцист, «глазки», обонятельные ямки, щупальца). Вслед за этим на переднем конце тела появляется скопление нервной ткани, из которой формируется мозговой или церебральный ганглий (рис. 10, 3
). У клеток церебрального
Рис. 10. Схема строения ортогональной нервной системы ресничного червя (передний конец):
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4
,
5
).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей.
Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является
ганглий —
парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает
нейропиль —
переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
Рис. 11. Схема строения узловой нервной системы насекомого:
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
нервных стволов, которые называются коннективами.
На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих
, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
Рис. 12. Схема строения головного мозга насекомого (пчела). Левая половина — его сечение:
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг
членистоногих состоит из трех отделов: переднего —
протоцеребрума
, среднего —
дейтоцеребрума
и заднего —
тритоцеребрума.
Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки.
Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
Рис. 13. Схема строения ганглионарной нервной системы пластинчатожаберного моллюска (беззубка):
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
Нервная система моллюсков
также имеет
ганглионарное строение
(рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг.
От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Вернуться к оглавлению
© 2000- NIV
Нейронауки
Современная наука о нервной системе объединяет многие научные дисциплины: наряду с классическими нейроанатомией, неврологией и нейрофизиологией, важный вклад в изучение нервной системы вносят молекулярная биология и генетика, химия, кибернетика и ряд других наук. Такой междисциплинарный подход к изучению нервной системы нашёл отражение в термине — нейронаука (neuroscience). В русскоязычной научной литературе в качестве синонима часто используется термин «нейробиология». Одной из основных целей нейронауки является понимание процессов, происходящих как на уровне отдельных нейронов, так и нейронных сетей, итогом которых являются различные психические процессы: мышление, эмоции, сознание. В соответствии с этой задачей изучение нервной системы ведется на разных уровнях организации, начиная с молекулярного и заканчивая изучением сознания, творческих способностей и социального поведения.
Профессиональные сообщества и журналы
Общество нейронаук (SfN, the Society for Neuroscience)[4] — крупнейшая некоммерческая международная организация, объединяющая более 38 тыс. учёных и врачей, занимающихся изучением мозга и нервной системы. Общество было основано в 1969 году, штаб-квартира находится в Вашингтоне. Основной его целью является обмен научной информацией между учёными. С этой целью ежегодно проводится международная конференция в различных городах США и издается Журнал нейронаук (The Journal of Neuroscience)[5]. Общество ведет просветительскую и образовательную работу.
Федерация европейских обществ нейронаук (FENS, the Federation of European Neuroscience Societies)[6] объединяет большое количество профессиональных обществ из европейских стран, в том числе и из России. Федерация была основана в 1998 году и является партнером американского общества нейронаук (SfN). Федерация проводит международную конференцию в разных европейских городах раз в 2 года и выпускает Европейский журнал нейронаук (European Journal of Neuroscience)[7].
Примечания
- Благодаря тому, что тело этих животных состоит всегда из одного и того же набора клеток.
- White J. G., Southgate E., Thomson J. N., Brenner S.
The structure of the nervous system of the nematode
Caenorhabditis elegans
//
Phil. Trans. Royal Soc. London
,
314
, 1986. — P. 1—340. - Медников, 1994, с. 363.
- Society for Neuroscience -. www.sfn.org. Проверено 22 ноября 2015.
- The Journal of Neuroscience. www.jneurosci.org. Проверено 22 ноября 2015.
- Fens Home Page (англ.). FENS.org. Проверено 22 ноября 2015.
- European Journal of Neuroscience.
- Harriet Cole : Drexel’s Longest-Serving Employee.