Эти пути можно подразделить на две группы:
1) главный двигательный, или пирамидный, путь, tractus pyramidalis (корково-ядерный и корково-спинномозговые пути), несет импульсы произвольных движений из коры головного мозга к скелетным мышцам головы, шеи, туловища, конечностей через соответствующие двигательные ядра головного и спинного мозга;
2) экстрапирамидные двигательные пути, tractus rubrospindlis, tractus ves—tibulospinalis и др., передают импульсы от подкорковых центров к двигательным ядрам черепных и спинномозговых нервов, а затем к мышцам.
Пирамидный путь
tractus pyramidalis, относится система волокон, по которым двигательные импульсы из коры большого мозга, из предцентральной извилины, от гиганто-пирамидальных нейронов направляются к двигательным ядрам черепных нервов и передним рогам спинного мозга, а от них — к скелетным мышцам.
Пирамидный путь подразделяют на три части:
1) корково-ядерный — к ядрам черепных нервов;
2) латеральный корково-спинномозговой (пирамидный) — к ядрам передних рогов спинного мозга;
3) передний корково-спинномозговой (пирамидный)—также к передним рогам спинного мозга.
Корково-ядерный путь
tractus corticonuclearis, представляет собой пучок отростков гигантопирамидальных нейронов, которые из коры нижней трети предцентральной извилины спускаются к внутренней капсуле и проходят через ее колено. Далее волокна корково-ядерного пути идут в основании ножки мозга, образуя медиальную часть пирамидных путей. Корково-спинномозговые, а также корковоядерный пути занимают средние основания ножки мозга. Начиная со среднего мозга и далее, в мосту и продолговатом мозге волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов: III и IV — в среднем мозге; V, VI, VII — в мосту; IX, X, XI, XII—в продолговатом мозге. В этих ядрах корково-ядерный (пирамидный) путь заканчивается, составляющие его волокна образуют синапсы с двигательными клетками этих ядер.
Центральный и периферический параличи, локализация поражения, клинические проявления.
Стр 1 из 13Следующая ⇒
Кортико-мышечный путь, локализация центральных и периферических нейронов в коре, стволе головного мозга, спинном мозге.
Кортико-мышечный путь –двухнейронный путь, соединяющий кору больших полушарий мозга со скелетной (поперечно-полосатой) мускулатурой.
Пирамидная система – совокупность всех центральных двигательных нейронов.
Этот путь состоит из двух нейронов: центральный (верхний) двигательный нейрон; периферический (нижний) двигательный нейрон.
Центральный нейрон нах-ся в области следующих извилин: верхний отдел прецентральной извилины (движение нижней конечности); средний отдел прецентральной извилины (движение верхней конечности); нижний отдел прецентральной извилины (движения головы, лица, языка, глотки, гортани); задний отдел верхней лобной извилины (движения туловища); задний отдел средней лобной извилины (поворот головы и глаз в противоположную сторону); Нейроны расположены, в основном, в 4-м цитоархитектоническом поле (по Бродману), а также в соседних полях. Центральный нейрон берет начало от больших пирамидных клеток Беца, расположенных в пятом кортикальном слое.
Импульсы пирамидных клеток идут по двум путям: корково-ядерный(оканчивается в ядрах черепных нервов); корково-спинномозговой (оканчивается в переднем роге спинного мозга). Аксоны всех этих нервных клеток идут вниз и кнутри, образуя в белом веществе лучистый венец. Далее они переходят во внутреннюю капсулу (корково-ядерный путь – колено внутренней капсулы;корково-спинномозговой путь – передние две трети заднего бедра внутренней капсулы). После выхода из внутренней капсулы, оба пучка переходят в ножки мозга (корково-ядерный путь расположен медиально, корково-спинномозговой – латерально). В мосту мозга тракт проходит в основании и делится на отдельные пучки. В пределах ствола мозга одна часть корково-ядерных волокон переходит на противоположную сторону, обеспечивается двусторонняя иннервация для глазодвигательных, жевательных мышц, для верхних мимических мышц, для мышц глотки и гортани. Другая часть корково-ядерных волокон переходит полностью на противоположную сторону, мышцы нижней половины лица и мышцы языка получают иннервацию только от коры противоположного полушария. Центральные нейроны корково-ядерного пути образуют синапсы с периферическими нейронами в ядрах черепных нервов: (тройничный, лицевой, языкоглоточный, блуждающий, подъязычный). Аксоны периферических нейронов корково-ядерного пути, составляют двигательные волокна черепных нервов. Волокна корково-спинномозгового пути сближаются и на поверхности продолговатого мозга образуют два валика – пирамиды. 80-85% волокон корково-спинномозгового пути, на границе продолговатого и спинного мозга, переходят на противоположную сторону и образуют – латеральный пирамидный пучок, остальная часть волокон спускаются неперекрещенными в передних канатиках СМ – передний пирамидный путь.Неперекрещенные волокна перекрещиваются на сегментарном уровне через переднюю комиссуру СМ. В шейной и грудной частях СМ некоторые волокна соединяются с клетками переднего рога своей стороны, так что мышцы шеи и туловища получают кортикальную иннервацию с обоих сторон.
Клетки периферических нейронов корково-спинномозгового пути расположены в передних рогах СМ и их аксоны входят в состав передних двигательных корешков и составляют двигательные волокна периферических спинномозговых нервов.
Центральный и периферический параличи, локализация поражения, клинические проявления.
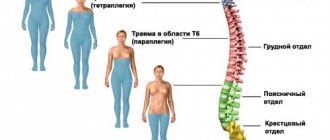
Параличи и парезы – утрата или ограничение двигательной ф-ии, хар-ся отсутствием (параличи) или снижением (парезы) мышечной силы, в результате чего выполнение движений невозможно или затруднено. В зависимости от места и объема поражения расстройства движения могут наблюдаться: моноплегия; параплегия; гемиплегия; диплегия; триплегия; тетраплегия.
Центральный паралич возникает при поражении центральных мотонейронов, на любом уровне корково-спинномозгового пути.
Симптомы центрального паралича:
— снижение силы в сочетании с утратой тонких движений; — гипертонус (спастическое повышение тонуса); — повышение проприоцептивных рефлексов с клонусом или без него; — снижение или утрата экстероцептивных рефлексов (брюшных, кремастерных, подошвенных); — появление патологических рефлексов; — защитные рефлексы; — патологические содружественные движения (синкинезии); — отсутствие реакции перерождения.
Спастический паралич – повышенный мышечный тонус. Клонусы –крайняя степень повышения глубоких рефлексов и расширения их рефлекторной зоны.
Патологические рефлексы: На нижней конечности патологические рефлексы подразделяют на разгибательные и сгибательные.
Разгибательные: рефлекс Бабинского – в ответ на штриховое раздражение стопы – разгибание I пальца, веерообразное расхождение всех остальных; реф-с Оппенгейма – разгибание I пальца стопы в ответ на проведение с нажимом подушечкой I пальца кисти по передней пов-ти голени; реф-с Гордона –разгибание I пальца или всей нижней конечности при сдавлении кистью икроножной мышцы; реф-с Шевера– разгибание большого пальца в ответ на сдавление пяточного сухожилия;реф-с Чеддона –разгибание I п стопы при штриховом раздражении кожи лодыжки в направлении от пятки к тылу стопы; реф-с Гроссмана –разгибание Iп стопы при сдавлении дистальной фаланги V п стопы.
Сгибательные: реф-с Россолимо – при отрывистых ударах по дистальным фалангам пальцев, происходит быстрое подошвенное сгибание всех пальцев стопы; реф-с Бехтерева-Менделя – при постукивании по тылу стопы в обл 3-4 плюсневых костей — быстрое сгибание 2-5 пальцев; реф-с Жуковского-Корнилова –подошвенное сгибание II-IV п стопы при ударе по подошвенной стороне стопы ближе к пальцам; верхний реф-с Россолимо –сгибание дистальной фаланги Iп кисти при коротком ударе по кончикам 2-5 пальцев; реф-с Россолимо-Вендеровича –удар наносится по дистальным фалангам слегка согнутых 4-5п; реф-с Бехтерева –кивательное движение 2-5п при ударе по тылу кисти в обл 2-4 пястных костей; реф-с Жуковского –сгибание 2-5п в ответ на удар по ладонной пов-ти кисти в обл 3-4 пястных костей; реф-с Гоффманна –сгиб-е движение пальцев в ответ на щипковое раздражение ногтевой пластинки 3 п свисающей руки; реф-с Клиппеля-Вейля –сгиб-е 1 кисти при пассивном разгибании 2-5п; симптом Якобсона-Ласка –ладонное сгиб-е пальцев кисти при ударе по лат части запястно-лучевого сочленения.
Патологические рефлексы осуществляемые ротовой мускулатурой: назолабиальный рефлекс – при постукивании по спинке носа пр-т сокращение круговой мышцы рта (вытягивание губ вперед); хоботковый рефлекс – при постукивании по верхней/нижней губе происходит вытягивание губ вперед.
Защитные рефлексы: реф-с Бехтерева-Мари-Фуа – при усиленном пассивном подошвенном сгибании пальцев или стопы происходит сгибание в тазобедренном, коленном и голеностопном суставах; реф-с Ремака – при штриховом раздражении кожи верхней трети передней области бедра происходят подошвенное сгибание стопы, 1-3 пальцев стопы и разгибание нижней конечности в коленном суставе; укоротительный реф-с Давиденкова –при штриховом раздражении стопы парализованной конечности или попытке произвести подошвенное сгибание стопы происходит сгибание в тазобедренном и коленном суставах и разгибание в голеностопном.
Синкинезия – рефлекторное содружественное движение конечности (или другой части тела), сопутствующее произвольному движению другой конечности (части тела). Если больной по заданию оказывает здоровой рукой сопротивление разгибанию в локтевом суставе, производимому исследующим, или сильно пожимает ему здоровой кистью руку, то в парализованной руке происходит сопутствующее рефлекторное сгибание.
Периферический паралич возникает при поражении второго нейрона двухнейронного двигательного пути, принимающего участие в иннервации мышцы, т.е. клеток передних рогов СМ, передних корешков и двигательных волокон спинномозговых и черепных периферических нервов. Хар-ся: утратой рефлексов; атонией или гипотонией мышц; атрофией мышц; реакцией перерождения. Реакция перерождения– характерные для периферического паралича изменения электрическое реакции пораженных нервов и мышц. При реакции перерождения нерв не проводит ток к мышце т.к. нерв поврежден, сама мышца денервирована и не способна сокращаться при непосредственном ее раздражении током. При периферических параличах в атрофических мышцах могут наблюдаться фибриллярные подергивания – быстрые сокращения отдельных мышечных волокон или их пучков.
| Уровень поражения | Симптоматика |
| Прецентральная извилина лобной доли: А) очаг деструкции Б) очаг возбуждения | А) контрлатеральный центральный или спастический монопарез. Б) эпилептические судорожные припадки (фокальные, моторные припадки тонические, тонико-клонические, клонические). |
| Внутренняя капсула | Контрлат спастический паралич, гемианестезия, гемианопсия, гемиплегия, центр парез 7, 12 пар. |
| Ствол, поражение периферических мотонейронов и кабелей от мотонейронов (фибрилляция/фасцикуляция). | Вялый контрлат паралич мышц, кот-е иннервируются этими ядрами. Перемежающийся (альтернирующий) синдром, чередуются стороны, где набл-ся поражение (3 этажа). Синдром Вебера – при поражении ножки мозга на стороне очага – поражение глазодвиг нерва, а противоположной – спастическая гемиплегия или гемипарез. Синдром Джексона –синдром периферич паралича мышц половины языка на стороне патолог очага в сочетании с гемипарезом или гемиплегией по центральному типу на противоположной части. Синдром Мийара-Гублера –периф паралич мимических мышц на стороне очага и гемиплегия на протиполож. |
| Верхний шейный сегмент С1 – С4 | Центральный тетрапарез (плегия), парез диафрагмы, наруш вегетативных ф-ий тазовых орг по центральному типу, утрата всех видов чув-ти ниже паралича с уровня затылка. |
| Шейное утолщение С5 – Th2: А) передние рога Б) боковые столбы | А) комбинированная тетраплегия: комбинированная верхняя вялая параплегия, нижняя спастическая параплегия Б) комбинированное: левосторонний верхний вялый монопарез, сниж чув-ти по проводниковому типу, левосторонний нижний спастический монопарез, наруш ф-ий тазовых орг. Синдром Горнера: миоз, птоз, энофтальм. Нарушение симпатической иннервации зрачка – (в боковых рогах – цилиоспинальный центр С8 – Th2). |
| Верхний грудной отдел | Неподвижная грудная клетка, вялый паралич наружных межреберных мышц, центральный паралич нижних конечностей (спастический парапарез), наруш ф-ии тазовых орг по проводниковому типу. |
| Нижний грудной отдел Th7 – Th12 | Центральный паралич ниж конечностей (параплегия), наруш ф-ий тазовых орг по центральному типу, проводниковое наруш чув-ти. |
| Поясничное утолщение (в передних рогах) | Нижний вялый парапарез, наруш ф-ий тазовых орг по центральному типу, проводниковое наруш чув-ти. |
| Эпиконус L4 – S2 | Выпадение, вялый паралич мышц задней пов-ти бедра, голени, ягодиц на стороне поражения. |
| Конус S3 – S5 | Утрата чув-ти в обл промежности, истинное недержание, наруш ф-ии тазовых орг по периферическому типу. |
| Конский хвост | Боли! Вялые ассиметричные парезы ниж конечностей, наруш ф-ии тазовых орг по периферическому типу. |
| Спинной мозг: А)передние рога Б) передние корешки | А) фибриллярные подергивания Б) фасцикулярные подергивания |
| Периферический нерв | Вялые параличи, парезы. |
1Следующая ⇒
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
Латеральный и передний корково-спинномозговые (пирамидные) пути
tractus corticospinales (pyramiddles) lateralis et ventralis, также начинаются от гигантопирамидальных нейронов предцентральной извилины. Аксоны этих клеток направляются к внутренней капсуле, проходят через переднюю часть ее задней ножки, спускаются в основание ножки мозга. Далее корково-спинномозговые волокна спускаются в переднюю часть (основание) моста и выходят в продолговатый мозг, где образуют пирамиды. В нижней части продолговатого мозга часть волокон переходит на противоположную сторону и продолжается в боковой канатик спинного мозга. Эта часть пирамидных путей, участвующая в образовании перекреста пирамид (моторный перекрест), получила название латерального корково-спинномозгового (пирамидного) пути. Те волокна корково-спинномозгового пути, которые не участвуют в образовании перекреста пирамид и не переходят на противоположную сторону, продолжают свой путь вниз в составе переднего канатика спинного мозга. Эти волокна составляют передний корково-спинномозговой (пирамидный) путь. Затем эти волокна также переходят на противоположную сторону, но через белую спайку спинного мозга и заканчиваются на двигательных клетках переднего рога противоположной стороны спинного мозга.
Корково-спинномозговой (пирамидный) и корково-ядерный пути
осуществляют сознательную произвольную регуляцию движений. Эти пути составляет цепочка из двух нейронов — центрального
и
периферического.
Центральными нейронами называют
клетки коркового ядра двигательного анализатора,
которые заканчиваются в двигательных ядрах черепных нервов (fibrae corticonuclearis) и в двигательных ядрах передних рогов спинного мозга
(tractus corticospinalis).
Периферическими нейронами являются
клетки двигательных ядер черепных нервов
и
двигательных ядер спинного мозга,
отростки которых в составе нервных стволов заканчиваются в соответствующих мышцах двигательными окончаниями. Пирамидный путь проводит импульсы от коркового ядра к активной части двигательного аппарата — мышцам. Эти импульсы возникают в коре как результат анализа и синтеза раздражений, поступивших в нее по проприоцептивному пути, а также по другим путям (зрительным, вестибулярным, слуховым, кожным). Основную массу волокон составляют аксоны пирамидных клеток предцентральной извилины.
В средних отделах моста от двигательного пути отделяется пучок волокон к двигательному ядру тройничного нерва своей и противоположной сторон. От клеток этих ядер возникают двигательные волокна тройничного нерва, которые в составе нижнечелюстного нерва идут к соответствующим мышцам и заканчиваются в них двигательными окончаниями. В нижнем отделе моста от двигательного пути отделяются волокна к двигательным ядрам лицевого нерва своей и противоположной сторон, после чего пирамидный путь переходит в продолговатый мозг, где снова принимает вид компактного пучка. В верхнем отделе продолговатого мозга от пирамидного пути отходят волокна к двигательным ядрам блуждающего, языкогло-точного и подъязычного нервов. После отхождения этих пучков пирамидный путь состоит только из волокон, соединяющих кору с ядрами передних рогов спинного мозга. Большая часть этих волокон совершает перекрест в нижнем отделе продолговатого мозга (decmsatio pyramidum)
и переходит в боковой канатик спинного мозга, где образует
латеральный(пирамидный) корково-спинальный путь,tractus corticospinalis lateralis.
Волокна этого пути заканчиваются в двигательных ядрах передних рогов спинного мозга. Меньшая часть волокон в области продолговатого мозга перекреста не образует. Эти волокна в спинном мозге составляет
передний корково-спинальный путь,tractus corticospinalis ventralis,
часть волокон которого заканчи-
вается в двигательных ядрах спинного мозга своей стороны, а другая часть волокон через переднюю белую спайку совершает перекрест и заканчивается в таких же ядрах противоположной стороны. Передний пучок прослеживается только в шейных и верхних грудных сегментах спинного мозга.
Таким образом, пирамидный путь осуществляет преимущественно перекрестную иннервацию мускулатуры тела, и при одностороннем поражении центральных нейронов любого отдела наступает паралич мышц на противоположной очагу поражения стороне. Такая односторонняя иннервация характерна не для всех мышечных групп. Большая часть мышц, а именно мышцы глазного яблока, жевательные, мимические мышцы верхней части лица, мышцы глотки, гортани, шеи, туловища и промежности, получают так называемую двустороннюю иннервацию, т. е. от центральных нейронов как своей, так и противоположной стороны. Это в известной степени обеспечивает сохранение иннервации этих мышц при одностороннем поражении центральных нейронов.
Односторонняя иннервация у мышц конечностей, языка и мимических мышц нижней половины лица. В связи с этим при одностороннем поражении центральных нейронов этих мышц наступает полная утрата способности к движению, т.е. паралич.
2. Проводящие пути экстрапирамидной системыпроходят к передним рогам спинного мозга вне пирамидных путей, через покрышку и ретикулярную формацию мозгового ствола. К экстрапирамидной системе относят следующие образования: хвостатое и чечевицеобразное ядро, таламус, черное вещество, красное ядро, образования гипоталамуса, мозжечок, нижнюю оливу и ядра ретикулярной формации мозгового ствола. По сравнению с пирамидой экстрапирамидная система является филогенетически более старым образованием, функция которого заключается в осуществлении сложных безусловнорефлек-торных двигательных актов, основанных на опыте вида, в то время как двигательная сознательная корковая деятельность основана на условных рефлексах, закрепляющих индивидуальный опыт.
Экстрапирамидная система обеспечивает автоматические, без участия сознания, регуляцию и координацию тонуса мышечной ткани, осуществляет сокращение или расслабление определенных групп мышц, обеспечивает готовность двигательного аппарата к выполнению сложных, точных, целенаправленных действий. Поражения экстрапирамидной системы проявляются главным образом изменениями мышечного тонуса и двигательной активности.
ТИПОВЫЕ ОСОБЕННОСТИ СТРОЕНИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Центральная нервная система у разных людей имеет некоторые специфические структурно-функциональные отличия. Эти отличия касаются формы мозга и его отдельных образований, величины, массы и других показателей.
Так, масса головного мозга человека, как правило, составляет от 1100 до 2300 г. Следует отметить, что большая масса мозга (в пределах нормы) не свидетельствует о высоком интеллекте, и наоборот. Между формой головного мозга и формой черепа наблюдается определенное соответствие. Долихоце-фалической, мезоцефалической и брахицефалической формам черепа соответствует определенное положение его отделов, образований, борозд и т. д. Это обстоятельство имеет важное практическое значение в связи с возможностью проекций на наружные покровы головы различных ядер, борозд и извилин.
ОБОЛОЧКИ ГОЛОВНОГО МОЗГА
Твердая оболочка головного мозга, dura mater encephali,
прилегает к внутренней поверхности костей черепа и плотно сращена с ней в области основания и швов. Рыхлое соединение оболочки с костями свода черепа послужило основанием для выделения очень узкого
эпидурального пространства,
которое пронизано соединительнотканными волокнами, сосудами и нервами. К гладкой внутренней поверхности твердой оболочки прилегает
паутинная оболочка
головного мозга. Узкую щель между ними условно называют
субдуральным пространством.
В некоторых местах твердая оболочка образует отроги (отростки), которые проникают между различными отделами головного мозга. Так, в продольной щели полушарий расположен серповидной формы отросток — серп большого мозга,falx cerebri.
Верхний утолщенный край серпа большого мозга, образованный двумя расходящимися пластинками твердой оболочки, примыкает к сагиттальной борозде черепа от петушиного гребня решетчатой кости до внутреннего затылочного выступа (рис. 213).
Намет мозжечка,tentorium cerebelli,
представляет собой широкую складку твердой мозговой оболочки, которая в виде двускатной крыши располагается в поперечной борозде мозга, отделяя содержимое задней черепной ямки от затылочных долей. Заднелатеральный утолщенный край мозжечкового намета сращен с бороздой поперечного синуса и верхней гранью пирамиды височной кости. Свободный край имеет округлую форму. Он натянут между задним концом свободного края
Рис. 213.Оболочки головного мозга. 1 _ грануляция паутинной оболочки; 2 — эмиссарная вена; 3 — вена губчатого вещества кости; 4 — губчатое вещество кости; 5 — твердая оболочка; 6 — перекладины паутинной оболочки; 7 — околососудистое пространство; 8 — подпау-тинное пространство; 9 — сосудистая оболочка; 10 — паутинная оболочка; 11 — серп большого мозга; 12 — верхний сагиттальный синус; 13 — кора мозга; 14 — ветвь артерии мозга; 15,16 — вены мозга.
мозгового серпа и задним наклонным отростком клиновидной кости. Свободный край ограничивает вырезку намета мозжечка, incisura tentorii,
через которую сообщаются задняя и средняя ямки черепа. В области этой вырезки расположены ножки мозга. В задней ямке черепа, между полушариями мозжечка, лежит
серп мозжечка,falx cerebelli.
Диафрагма турецкого седла,diaphragma sellae,
натянута над турецким седлом. В середине диафрагмы имеется узкое отверстие, через которое проходит воронка к гипофизу. На верхушке пирамиды височной кости отростки твердой оболочки образуют тройничную полость,
cavum trigeminale;
в ней расположен тройничный узел.
В толще твердой оболочки проходят артерии, вены, нервные волокна и их окончания, а также особые каналы — венозные синусы (пазухи).
Венозные синусы
представляют собой пространства между расщепившимися листками твердой оболочки головного мозга. Большая часть их расположена пристеночно. В венозные синусы поступает кровь из вен головного мозга и затем выводится в экстракраниальные венозные образования, главным образом во внутренние яремные вены.
Паутинная оболочка головного мозга, arachnoidea mater encephali,
в виде тонкого прозрачного бессосудистого листа по-
крывает головной мозг и книзу переходит в паутинную оболочку спинного мозга. С твердой оболочкой головного мозга паутинная оболочка прочно сращена посредством особых структур — грануляций паутинной оболочки, а
также в местах вступления вен в венозные синусы.
Паутинная и мягкая оболочки ограничивают заполненное спинномозговой (церебральной) жидкостью подпаутинное пространство.
Это пространство пронизано многочисленными соединительнотканными волокнами, которые соединяют обе оболочки и осуществляют фиксацию расположенных здесь кровеносных сосудов. Иные соотношения этих оболочек в области различных углублений мозга. Мягкая оболочка покрывает мозговую поверхность в глубине борозд, щелей и ямок, а паутинная, не следуя за ней в глубину борозд и щелей, как бы мостиками перекидывается с одной возвышенности извилины на другую: образуя различной величины подпаутинные пространства. Самые крупные из этих вместилищ носят название
подпаутинных цистерн:
1) мозжечково-мозговая цистерна, cistema cerebellomedullaris,
находится между мозжечком и задней поверхностью продолговатого мозга;
2) межножковая цистерна, cistema interpeduncularis,
расположена между ножками мозга;
3) цистерна перекреста, cistema chiasmatis,
находится впереди зрительного перекреста;
4) цистерна латеральной ямки большого мозга, cistema fossae lateralis cerebri,
соответствует одноименной ямке и борозде.
Субарахноидальное пространство является единым для головного и спинного мозга. Образующаяся в желудочках головного мозга спинномозговая жидкость поступает в это пространство через отверстия IV желудочка, а отводится в венозную систему посредством грануляций паутинной оболочки.
Мягкая оболочка головного мозга, pia mater encephali,
плотно прилегает к мозговому веществу как на свободной поверхности мозга, так и в глубине борозд. Сосудистые сплетения желудочков осуществляют секрецию спинномозговой жидкости. В толще мягкой оболочки расположены сети кровеносных сосудов и нервы.
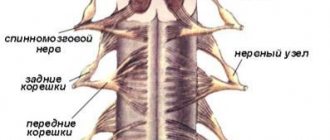
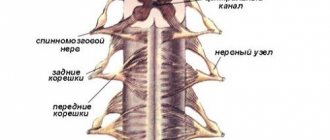
СПИННОМОЗГОВЫЕ НЕРВЫ
Спинномозговые нервы,
ил.
spindles,
располагаются в правильном порядке (невромеры), соответствуя миотомам (миомерам) туловища и чередуясь с сегментами позвоночного столба; каждому нерву соответствует относящийся к нему участок кожи (дерматом)
У человека имеетс31 пара З спинномозговых нервов, а именно: 8пар шейнь 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых (см. рис. 267). Каждый спинномозговой нерв отходит от спинного мозга двумя корешками: з а д н и м (чувствительным) и п е р е д н им (двигательным); оба корешка соединяются один ствол, trucnus n. spinalis выходящий из позвоночного канала (через межпозвоночное отверстие Вблизи и несколько кнаружи от места соедитения задний корешок образует узел, ganglion spindle,
в котором передний двигательный корешок не принимает участия. Благодаря соединению обоих корешков спинномозговые нервы явля-ются смешанными нервами: они содержат чувствительные (яфтрерёнтнмр) волокна от клеток спинномозговых узлов, двигательные (эфферентные) волокна от клеток переднего рога, а также вегетативные волокна от клеток боковых рогов, выходящие из спинного мозга в составе переднего корёшкги (рис. 308).
Вегетативные волокна имеются и в заднем корешке. Вегетативные волокна, попадающие через корешки в анимальные нервы, обеспечивают в соме такие процессы, как трофика сосудодвигательные реакции и т. п.
У круглоротых (миноги) оба корешка продолжаются в отдельные нервы двигательные и чувствительные. В дальнейшем ходе эволюции, начиная с поперечноротых рыб корешки сближаются и сливаются, так что раздельный ход сохраняется только для корешков, а нервы становятся смешанными. Каждый спинномозговой нерв при выходе из мезенхимы|рмттрг.нрчного отверстия делится соответственно двум частям миптомя. (дорсальной и вентральной) на две ветви:
1) заднюю, rumusi dprsgljs,для развивающейся из дорсальной части миото-ма аутохтонной»мускулатуры спины и покрывающей ее кожи;
2) переднюю, ramus ventralis,для вентральной стенки туловища и конеч-ностей, развивающихся из вентральных частей миотомов.
Кроме того, от спинномозгового нерва отходят еще два рода ветвей:
3) для иннервации внутренностей и сосудов — соединительные ветви к симпатическому стволу, rr. communicantes;
4) для иннервации оболочек спинного мозга — г. menfngeus,идущая обратно через межпозвоночное отверстие.
Преддверно-спинномозговой путь
Важным звеном в координации двигательных функций тела человека является преддверно-спинномозговой путь, tractus ves—tibulospinalis. Он связывает ядра вестибулярного аппарата с передними рогами спинного мозга и обеспечивает установочные реакции тела при нарушении равновесия. В образовании пред-дверно-спинномозгового пути принимают участие аксоны клеток латерального вестибулярного ядра, а также нижнего вестибулярного ядра преддверно-улиткового нерва. Эти волокна спускаются в латеральной части переднего канатика спинного мозга (на границе с боковым) и заканчиваются на двигательных клетках передних рогов спинного мозга.
Анатомия проводящих путей нервной системы
Содержание
После этого волокна tractus tectospinalis через ствол «направляются» к сегментам спинного мозга. В покрышке моста этот путь занимает дорсомедиальное положение, несколько вентральнее продольных пучков. Сходная топография наблюдается и в продолговатом мозге, где tractus tectospinalis находится вентральнее медиального продольного пучка и постепенно смещается вентрально, приближаясь к дорсальной границе пирамид. В спинном мозге он находится в медиальной части переднего канатика. Постепенно крышеспинномозговой путь истончается, так как часть его волокон заканчивается на мотонейронах двигательных ядер черепных нервов в стволе (крышеядерный пучок, fasciculus tectonuclearis) и в вышележащих сегментах спинного мозга. Здесь, через интернейроны, волокна tractus tectospinalis влияют на альфа-малые мотонейроны двигательных ядер передних рогов.
Мотонейроны ствола и спинного мозга по своим аксонам передают влияние от интеграционного центра крыши среднего мозга через черепные и спинномозговые нервы к иннервируемым скелетным мышцам.
Поражение tractus tectospinalis приводит к утрате стартовых рефлексов на внезапные световые, звуковые, обонятельные и тактильные воздействия.
Ретикулоспинномозговой путь
Этот путь считают наиболее филогенетически старым и неспецифическим. При этом под названием «tractus reticulospinalis» понимают совокупность эфферентных волокон, начинающихся от различных центров ретикулярной формации и имеющих функциональные и топографические особенности. В упрощенном виде ретикулоспинномозговой путь может быть изображен без перекреста, без интернейронов, без указания конкретного ядра, от которого он начинается, и в виде одиночной, а не множественной проекции (рис. 18).
Рис. 18. Ретикулоспинномозговые пути: 1 — ретикулярные ядра, 2 — ретикулоспинномозговой путь, 3 — двигательные ядра передних рогов спинного мозга, 4 — спинномозговые нервы
Следует учитывать, что из себя представляет ядро-мишень в спинном мозге: в случае анимальной рефлекторной дуги это двигательные ядра переднего рога, а в случае симпатической рефлекторной дуги — промежуточно-боковое ядро бокового рога.
Другими словами, существует несколько параллельных ретикулоспинномозговых путей.
Медиальный ретикулоспинномозговой путь (tractus reticulospinalis medialis) — самый мощный и протяженный из ретикулоспинномозговых путей. Он начинается из орального и каудального ретикулярных ядер моста и из ретикулярных ядер продолговатого мозга: гигантоклеточного и вентрального. В спинном мозге он простирается до крестцовых сегментов, постепенно истончаясь и посегментно заканчиваясь на дендритах гамма-мотонейронов передних рогов спинного мозга.
Латеральный ретикулоспинномозговой путь (tractus reticulospinalis lateralis) начинается из латерального ретикулярного ядра моста, расположенного около средней ножки мозжечка (regio parabrachialis). Этот путь частично перекрещенный, включает в свой состав аксоны ретикулярных нейронов дыхательного в спинной мозг, где располагается в боковом канатике рядом с боковым корково-спинномозговым путем. Tractus reticulospinalis lateralis оказывает активирующее влияние на малые альфа-мотонейроны передних рогов спинного мозга. Другая часть его волокон заканчивается на нейронах промежуточно-бокового ядра спинного мозга (центр симпатического отдела вегетативной нервной системы). Поэтому становится возможной регуляция органов «растительной жизни» со стороны ретикулярной формации.
Передний ретикулоспинномозговой путь (tractus reticulospinalis anterior) начинается из покрышечных ретикулярных ядер среднего мозга и моста и, располагаясь в передних канатиках спинного мозга, «достигает» десятого грудного сегмента. Этот путь заканчивается на мотонейронах передних рогов спинного мозга.
Для всех ретикулоспинномозговых путей характерна лучшая выраженность в шейных и верхнегрудных сегментах спинного мозга. Дистальнее влияние ретикулярной формации распространяется по проприоспинальным путям. Другими словами, ретикулоспинномозговым путям свойственна форма цепочки из нескольких последовательно расположенных нейронов (полисинаптическая организация). Другой особенностью является то, что ретикулоспинномозговые пути преимущественно неперекрещенные. Все эти пути имеют опосредованную связь с мотонейронами передних рогов, так как заканчиваются на дендритах интернейронов 7 и 8 пластин по Рекседу и уже через них влияют на мотонейроны. Эти влияния могут быть как тормозного, так и активирующего характера. В результате ретикулярная формация через свои ретикулоспинномозговые пути и спинномозговые нервы обеспечивает тонус скелетных мышц и выполнение сложных рефлекторных актов, требующих одновременного участия многих скелетных мышц или даже групп мышц (дыхательные, хватательные движения). Сходные отношения имеются между центрами ретикулярной формации и ядрами черепных нервов.
Преддверно-спинномозговой путь
Этот путь также относится к весьма древним в эволюционном плане проекциям, тесно связанным с вестибулярным анализатором. Tractus vestibulospinalis участвует в быстрой реакции организма на такое изменение положения тела в пространстве, которое приводит к нарушению равновесия. При этом происходят безусловно-рефлекторные телодвижения, приводящие к тому, что человек, поскользнувшись, падает на выставленные руки и не ударяется головой или туловищем.
Начинается этот путь из латерального вестибулярного ядра (ядра Дейтерса) (nucl. vestibularis lateralis), расположенного в покрышке моста недалеко от границы последнего с продолговатым мозгом (рис. 19).
Рис. 19. Преддверно-спинномозговой путь: 1 — вестибулярные ядра, 2 — преддверно-спинномозговой путь, 3 — двигательные ядра передних рогов спинного мозга, 4 — спинномозговые нервы
По данным ряда исследователей, в состав tractus vestibulospinalis входят также аксоны нейронов, тела которых расположены в нижнем вестибулярном ядре (ядре Роллера). Последнее расположено рядом с ядром Дейтерса, но несколько каудальнее. Ядро Дейтерса оказывает опосредованное влияние (в частности, через альфа-мотонейроны двигательных ядер передних рогов спинного мозга) на мышцы-разгибатели и тем самым является своеобразным антагонистом красного ядра. В продолговатом мозге преддверно-спинномозговой путь располагается дорсальнее и латеральнее пирамид, а в спинном мозге — на границе переднего и бокового канатиков (здесь он пронизан волокнами передних корешков спинномозговых нервов). Путь преимущественно неперекрещенный.
Оливоспинномозговой путь
Tractus olivospinalis участвует в безусловно-рефлекторном поддержании тонуса мышц шеи и в выполнении движений, призванных сохранять равновесие тела. Этот путь является относительно молодым в эволюционном плане, как и ядро оливы (nucleus olivaris) продолговатого мозга, от которого он начинается. На ядро оливы оказывают регулирующее влияние полушария мозжечка (кора и зубчатое ядро), красное ядро и кора лобной доли полушария большого мозга.
Аксоны нейронов nucl. olivaris в составе tractus olivospinalis достигают шестого шейного сегмента спинного мозга, посегментно заканчиваясь на альфа-мотонейронах двигательных ядер передних рогов на своей стороне тела (рис. 20).
Рис. 20. Оливоспинномозговой путь: 1 — ядра нижней оливы, 2 — оливоспинномозговой путь, 3 — двигательные ядра передних рогов спинного мозга, 4 — спинномозговые нервы, 5 — мышцы шеи
Аксоны этих мотонейронов в составе спинномозговых нервов достигают мышц шеи, которые и иннервируют. В спинном мозге оливоспинномозговой путь расположен в переднемедиальном отделе бокового канатика.
3.2. Пирамидные пути
Эти пути, в совокупности называемые еще «пирамидная система», участвуют в сознательном контроле функции скелетных мышц (стимулирование или торможение сокращения). В частности, возможно выполнение произвольных движений, характеризующихся сложностью и точностью. Пирамидная система состоит из двух путей: корково-спинномозгового (tractus corticospinalis) и корково-ядерного (tractus corticonuclearis). Свое название пирамидная система получила в связи с тем, что tractus corticospinalis «проходит» через пирамиды продолговатого мозга. Понятно, что название не слишком удачное, так как главным здесь является не топография, а функция.
Корково-спинномозговой путь
Этот путь проводит волевые двигательные импульсы, позволяющие управлять скелетными мышцами, иннервируемыми спинномозговыми нервами, т.е. мышцами конечностей, туловища и шеи. Корково-спинномозговой путь проводит также импульсы, способные тормозить активность мотонейронов передних рогов спинного мозга.
VK
Buffer
Корково-мостомозжечковому путь
Управление функциями мозжечка, участвующего в координации движений головы, туловища и конечностей и связанного в свою очередь с красными ядрами и вестибулярным аппаратом, осуществляется из коры большого мозга через мост по корково-мостомозжечковому пути, tractus corticopontocerebellaris. Этот проводящий путь состоит из двух нейронов. Тела клеток первого нейрона лежат в коре лобной, височной, теменной и затылочной долей. Клетки ядер моста с их отростками составляют второй нейрон корково-мостомозжечкового пути.