В спинном мозге рефлекторные кольца скелетных мышц взаимосвязаны с рефлекторными кольцами внутренних органов. Поэтому при раздражениях рецепторов афферентные нервные импульсы вызывают также возбуждение эфферентных нейронов, иннервирующих сердце, сосуды, железы и другие органы. При этом гормоны и другие химические продукты поступают в кровь и объектом их воздействия становится и центральная нервная система.
Следовательно, в спинном мозге имеются нейроны, вызывающие соматические (двигательные) и вегетативные рефлексы.
Основные свойства нервных центров спинного мозга имеют подчиненное значение, так как ведущая роль в объединении и регулировании всей деятельности организма в его взаимодействии с окружающим миром принадлежит высшему отделу нервной системы — большим полушариям головного мозга.
Определение возбудимости центров спинного мозга
Возбудимость спинного мозга определяется по длительности латентного периода рефлекса и по величине рефлекторной реакции. Латентный период рефлекса значительно больше, чем при раздражении нервно-мышечного препарата. Он складывается из времени проведения возбуждения по афферентному пути, по центральной нервной системе и по эфферентному пути. При детальных исследованиях учитывается также латентный период рецептора и эффектора.
Время прохождения возбуждения по центральной нервной системе или необходимое для перехода возбуждения с афферентных нейронов на эфферентные, называется центральным временем рефлекса, или редуцированным временем рефлекса. Его продолжительность зависит от количества нейронов и синапсов, участвующих в осуществлении данного рефлекса. Оно тем продолжительнее, чем больше нейронов и синапсов имеется на пути проведения возбуждения по центральной нервной системе, т. е. в рефлекторном кольце, так как проведение возбуждения задерживается в каждом нейроне и особенно в каждом синапсе.
Центральное время рефлекса при двунейронной рефлекторной «дуге», в которой имеется только один синапс, наименьшее. Продолжительность задержки возбуждения в каждом синапсе центральной нервной системы (или в мионевральном аппарате) минимально равна 0,3 мс (в каждом в среднем не более 0,5 мс).
Время сухожильных рефлексов самое короткое; например, коленного — 5,3 мс, из которых на проведение возбуждения по периферическим путям приходится 3,1 мс, а на центральное время — 2,2 мс. При повторении раздражения центральное время может уменьшиться до 1 и даже до 0,3 мс. Продолжительность центрального времени рефлекса наибольшая при слабых раздражениях. При сложных соматических рефлексах центральное время рефлекса увеличивается до 10-20 и даже до 500 мс, а при вегетативных рефлексах — до десятков секунд.
Изменения возбудимости
. Сразу же после проведения каждого нервного импульса повторные раздражения вызывают в нейронах спинного мозга абсолютную и относительную рефрактерную фазы, а затем наступает экзальтационная фаза. Эти фазы более продолжительны, чем в нервных волокнах.
После экзальтационной фазы возбудимость понижается. Ритм и сила рефлекторной реакции зависят от изменений возбудимости и лабильности нейронов. В связи с возникновением в одних нейронах спинного мозга возбуждения возбудимость их увеличивается, а в связи с возникновением в других нейронах нервной системы торможения возбудимость этих нейронов уменьшается. Следовательно, возбудимость нейронов спинного мозга постоянно изменяется в связи со сменой возбуждения и торможения и поэтому не существует постоянного порога рефлекторного раздражения.
Возбудимость изменяется в зависимости от притока импульсов, поступающих из рецепторов. Синапсы в течение многих дней могут находиться в состоянии резко пониженной возбудимости или, наоборот, в состоянии повышенной возбудимости, что в первом случае сопровождается отсутствием моторных рефлексов, а во втором — усилением рефлекторных движений.
Тест с ответами: “Строение и функции спинного мозга”
1. При поврежденном спинном мозге на уровне шейного отдела возможны повороты головы, можно говорить, совершать жевательные движения, так ли это: а) да + б) нет в) отчасти
2. У спинальной лягушки (без головного мозга) двигательные рефлексы сохраняются, так ли это: а) нет б) да + в) не изучено
3. Спинной мозг выполняет проводниковую функцию, так ли это: а) нет б) не изучено в) да +
4. При поврежденном спинном мозге на уровне шейного отдела любые движения становятся невозможными, так ли это: а) нет + б) да в) отчасти
5. Спинной мозг состоит из … сегмента: а) 28 б) 42 в) 31 +
6. Спинной мозг состоит из 42 сегментов, так ли это: а) да б) нет + в) отчасти
7. Толщина спинного мозга около 4 см, так ли это: а) да б) по-разному в) нет +
8. Спинной мозг выполняет рефлекторную функцию, так ли это: а) нет + б) да в) отчасти
9. Потеря чувствительности в определенных участках тела наблюдается при: а) повреждении передних рогов спинного мозга б) повреждении передних корешков спинномозговых нервов в) повреждении задних корешков спинномозговых нервов +
10. Паралич определенной группы мышц наблюдается при: а) разрушенном в спинном мозге б) повреждении задних корешков спинномозговых нервов + в) повреждении передних корешков спинномозговых нервов
11. От спинного мозга возбуждение проходит по: а) передним корешкам спинномозговых нервов + б) смешанным нервам к головному мозгу в) задним корешкам спинномозговых нервов
12. К спинному мозгу возбуждение проходит по: а) передним корешкам спинномозговых нервов б) задним корешкам спинномозговых нервов + в) смешанным нервам от головного мозга
13. Вставочные нейроны находятся: а) в передних рогах серого вещества спинного мозга б) в передних корешках спинномозговых нервов в) в задних рогах серого вещества спинного мозга +
14. Двигательные нейроны находятся в: а) узлах задних корешков спинномозговых нервов б) задних рогах серого вещества спинного мозга в) передних рогах серого вещества спинного мозга +
15. Чувствительные нейроны находятся в: а) узлах задних корешков спинномозговых нервов + б) передних корешках спинномозговых нервов в) задних рогах серого вещества спинного мозга
16. Белое вещество спинного мозга образовано отростками нейронов, так ли это: а) нет б) да + в) не изучено
17. В спинномозговом канале проходят нервы и кровеносные сосуды, так ли это: а) да б) не изучено в) нет +
18. Серое вещество спинного мозга образовано отростками нейронов, так ли это: а) да б) нет + в) не изучено
19. В каком отделе наблюдается утолщение спинного мозга: а) крестцовом б) грудном в) шейном +
20. В каком отделе наблюдается утолщение спинного мозга: а) грудном б) поясничном + в) крестцовом
21. Какая реакция (действие) организма с помощью рефлекторной дуги напрямую связана со спинным мозгом: а) чиханье + б) движение глаз в) приседания
22. Что такое восходящие пути: а) передача сигналов от головного в спинной мозг б) передача сигналов от спинного в головной мозг + в) передача сигналов от головного мозга к органам
23. Что такое серое вещество: а) скопление отростков нервных клеток б) скопление миелиновых волокон в) скопление нервных клеток +
24. Чем заполнено пространство между позвоночным каналом и спинным мозгом: а) белым веществом б) жировой тканью + в) серым веществом
25. Чем заполнено пространство между позвоночным каналом и спинным мозгом: а) серым веществом б) жировой тканью в) кровеносными сосудами +
26. Чем заполнено пространство между позвоночным каналом и спинным мозгом: а) белым веществом б) спинномозговой жидкостью + в) жировой тканью
27. В какой последовательности расположены оболочки спинного мозга (от центра к периферии): а) мягкая, паутинная, твёрдая + б) твёрдая, паутинная, мягкая в) твёрдая, мягкая, паутинная
28. Что такое спинномозговая нить: а) нервные волокна, отходящие от нейронов б) полый канал, проходящий через центр спинного мозга в) узкий отросток спинного мозга, срастающийся с надкостницей копчика +
29. Где заканчивается спинной мозг: а) на уровне первого поясничного позвонка + б) на уровне первого крестцового позвонка в) на последнем позвонке поясничного отдела
30. С каким отделом головного мозга соединён спинной мозг: а) передний мозг б) продолговатый мозг + в) промежуточный мозг
Координирующие механизмы спинного мозга
Согласованность, соподчиненность сокращений отдельных мышц или их групп, основанная на взаимодействии возбуждения и торможения в нервной системе, обозначается как координация.
Побуждение группы нейронов сопровождается изменением функционального состояния соседних нейронов, их торможением. По ному каждый рефлекс — результат взаимодействия возбуждении и торможения в центральной нервной системе, согласованной и соподчиненной деятельности многих нейронов. Именно благодаря координации нейронов, которая возникла в процессе фило- и онтогенетического развития, могут осуществляться тонкие движении, изолированные двигательные рефлексы вместо общих массовых рефлекторных движений, присущих организмам, у которые нервная система находится на низших ступенях фило- и онтогенеза. Координация характеризует не только сложные, но и самые простые рефлексы.
Участие спинного мозга в регуляции тонуса мышц
Регуляция тонуса мышц имеет рефлекторную природу и осуществляется посредством миотатических и позно-тонических рефлексов.
Миотатический рефлекс является рефлексом растяжения, который начинается с раздражения мышечного веретена. Практически в каждой мышце находятся мышечные рецепторы растяжения, называемые из-за своей формы «мышечными веретенами» (рис.2).
Мышечный рецептор состоит из поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой, и имеет форму веретена. Длина интрафузальных мышечных волокон равна 4 — 7 мм, толщина 15 — 30 мкм. Мышечные веретена располагаются параллельно скелетной мышце и своими концами крепятся к соединительнотканной оболочке пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5 — 1 мм.
Механизм регуляции тонуса мышц представлен на рисунке 2. Растяжение экстрафузальных мышечных волокон (при расслаблении поперечно-полосатых мышечных волокон) приводит к растяжению мышечного веретена. При растяжении мышечного веретена происходит растяжение ядерной сумки и возбуждение рецептора растяжения. Импульсы поступают в спинной мозг к α-мотонейронам. Возбуждение по аксонам α-мотонейронов вызывает сокращение экстрафузальнох мышечных волокон.
Рис.2. Схема регуляции мышечного тонуса.
1 – экстрафузальные волокна; 2 – мышечное веретено; 3 – интрафузальные
волокна; 4 – эфферентные нервные волокна; 5 – альфа-мотонейроны;
6 – гамма-мотонейроны
Гамма-петля:
Ряд воздействий от рецепторов мышц и сухожилий идет не только на α-мотонейроны, но и по коллатерали аксона на γ-мотонейроны: γ-мотонейроны возбуждаются → посылают импульсы к интрафузальным волокнам → интрафузальные волокна сокращаются → мышечное веретено растягивается → рецептор возбуждается → импульсы идут в спинной мозг → активация α-мотонейронов → сокращение экстрафузальных волокон.
Шейные позно-тонические рефлексы
спинного мозга направлены на поддержание позы. С их помощью регулируется тонус мышц. Эти рефлексы возникают с проприорецепторов мышц и фасций шеи. Переключение с этих рецепторов осуществляется на уровне шейного отдела спинного мозга и приводит к изменению тонуса мышц при изменении положения головы и шеи.
Сегментарный и межсегментарный принцип
Работы спинного мозга
Спинной мозг состоит из 31-32 сегментов. Каждому сегменту соответствуют две пары корешков (соответственно числу сегментов из спинного мозга выходит 31-32 пара передних двигательных корешков и входит в него 31-32 пары задних чувствительных корешков). При проекции сегментов спинного мозга на позвонки приходится учитывать несоответствие длины спинного мозга и позвоночника. В шейном отделе сегменты расположены на 1 позвонок выше, чем соответствующий им по счету позвонок; верхнегрудные – на 2, нижнегрудные – на 3 (пример: V шейный сегмент расположен на уровне IV шейного позвонка, V грудной – на уровне III грудного позвонка, XI грудной – на уровне VIII грудного позвонка и т.д.). В большом несоответствии находятся поясничные и крестцовые сегменты и позвонки: поясничные сегменты находятся на уровне X, XI, и XII грудных позвонков; крестцовые – XII грудного и I поясничного. Морфологических границ между сегментами спинного мозга нет, поэтому деление на сегменты является функциональным и определяется зоной распределения в сегменте волокон заднего корешка и зоной клеток, которые образуют передний корешок. Часть тела, иннервируется волокнами одного сегмента, называется метамер (участок кожи, иннервированные чувствительными волокнами сегмента, называется дерматом). Каждый сегмент иннервирует через свой метамер. Это сегментарный принцип работы спинного мозга. Межсегментарный принцип работы спинного мозга заключается в том, что каждый сегмент спинного мозга иннервирует не только свой метамер, но и метамеры выше- и нижележащих соседних сегментов. В итоге перекрытия нервных волокон каждый метамер тела иннервируется тремя сегментами и передает сигналы в 3 сегмента спинного мозга, что повышает надежность регуляторных механизмов.
Структурная организация и функции продолговатого мозга
Продолговатый мозг является продолжением спинного мозга и имеет длину около 25 мм. В отличие от спинного мозга его серое вещество расположено не в центре, а на периферии в виде ядер.
В продолговатом мозге располагаются ядра VIII-XII пар черепно-мозговых нервов через которые осуществляется регуляция чувствительных (сенсорных), двигательных (соматических) и вегетативных (парасимпатических) функций: чувствительные ядра преддверно-улиткового нерва (n.vestibulocochlearis, VIII), ядра языкоглоточного нерва (n. glossopharyngeus, IX), ядра блуждающего нерва (n. vagus, X), двигательное ядро добавочного нерва (n. accessorius, XI), двигательное ядро подъязычного нерва (n. hypoglossus, XII). Кроме того в РФ продолговатого мозга располагаются вестибулярные ядра (медиальное – ядро Швальбе, верхнее – ядро Бехтерева, латеральное – ядро Дейтерса и нижнее – ядро Роллера), к которым поступают импульсы от рецепторов полукружных каналов.
Функции продолговатого мозга
Продолговатый мозг выполняет сенсорные, проводниковые и рефлекторные функции.
— Сенсорные функции
осуществляются за счет поступления в продолговатый мозг афферентной информации от рецепторов вкуса, слуховых и вестибулярных рецепторов по чувствительным волокнам к ядрам соответствующих черепно-мозговых нервов и от рецепторов органов грудной и брюшной полости (см.выше). На уровне продолговатого мозга производится первичный анализ силы и качества раздражителя.
— Проводниковые функции
. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга. Здесь находятся перекресты нисходящих пирамидных путей и восходящего тракта Голля и Бурдаха.
— Рефлекторные функции
. В продолговатом мозге локализованы центры многочисленных рефлексов. Например, ряд защитных рефлексов:
рвотный рефлекс
(от рецепторов корня языка, глотки, желудка, кишечника, брюшины, вестибулярного аппарата → по волокнам языкоглоточного, блуждающего или вестибулярного нерва → в рвотный центр продолговатого мозга → по эфферентному волокну блуждающего нерва к пищеводу, желудку, кишечнику и через спинальные моторные центры к диафрагме и мышцам брюшной стенки);
кашлевой рефлекс
(от рецепторов гортани, трахеи и бронхов → по чувствительным волокнам блуждающего нерва → в кашлевой центр продолговатого мозга → эфферентные импульсы к спинальным центрам дыхательной мускулатуры);
рефлекс чиханья
(от рецепторов полости носа → в центр чиханья продолговатого мозга → эфферентные импульсы к спинальным центрам дыхательной мускулатуры), а также
рефлексы слезоотделения, смыкания век.
Также продолговатый мозг вместе с другими структурами ствола мозга участвует в осуществлении
рефлексов поддержания позы (вестибулярных позно-тонических рефлексов).
В продолговатом мозге кроме центров защитных рефлексов в области ретикулярной формации находятся жизненно важные центры – сосудодвигательный и дыхательный центры.
Особенности структурной организации ретикулярной формации
ствола мозга
Ретикулярная формация (РФ) располагается в сером веществе продолговатого мозга, среднего мозга, промежуточного мозга, частично спинного мозга и регулирует уровень активности коры головного мозга, мозжечка, таламуса, спинного мозга. РФ образована нейронами, которые имеют длинные маловетвящиеся дендриты и короткий хорошо ветвящийся аксон. Морфологическое строение ретикулярной формации очень напоминает сеточку, и именно по этому признаку немецкий анатом Отто Дейтерс назвал эту структуру «ретикулярная формация» (лат. reticulum — сеточка, formatio — образование) (1865). Нейроны РФ получают информацию от разных рецепторов. Это полимодальные нейроны, имеющие большие рецептивные поля, обладают спонтанной активностью, повышенной возбудимостью, высокой лабильностью (до 1000 имп/сек), высокой чувствительностью к метаболитам, гормонам, барбитуратам и другим фармакологическим препаратам.
Особенности иррадиации возбуждения в спинном мозге
Возбуждение, поступившее в спинной мозг по афферентному нейрону, распространяется на выше и ниже расположенные сегменты. Этот процесс распространения возбуждения называется иррадиацией.
Иррадиация возбуждения особенно ясно наступает вследствие увеличения силы раздражения. При слабом раздражении рецепторов наблюдается незначительный рефлекс, так как рефлекторное движение ограничивается небольшой группой мышц. При усилении раздражения рецепторов может наступить общая рефлекторная двигательная реакция. Величина рефлекса зависит от количеств двигательных нейронов, находящихся в состоянии возбуждения. При слабом раздражении рецепторов количество возбужденных эфферентных моторных нейронов невелико, а при усилении раздражении рецепторов, благодаря иррадиации возбуждения, и поэтому возбужденных нейронов все более возрастает, и поэтому начинают сокращаться все большие массы мускулатуры.
Иррадиация возбуждении по нервным центрам увеличивается также при возрастании продолжительности раздражения рецепторов, что приводит к возбуждению все большего количества нейронов. Торможение ограничивает иррадиацию возбуждения, пропуская возбуждение по определенным нервным путям.
Синергетические и антагонистические рефлексы
Различаются две группы мышц, участвующих в координированном движении: 1) синергисты и 2) антагонисты.
Одновременное совместное сокращение синергистов обеспечивает выполнение данного движения, например короткая, длинная и большая мышцы, сокращаясь одновременно, приводят бедро.
Совместное сокращение антагонистов вызывает движения противоположного характера и поэтому делает невозможным выполнение данного движения, например сгибатели и разгибатели, приводящие и отводящие мышцы. При выполнении других движений антагонистические мышцы могут стать синергистами при отведении и приведении или разгибатели обеих ног становятся синергистами во время стояния.
Реципрокные (сопряженные) отношения возбуждения и торможения в центральной нервной системе
Впервые Л. А. Спиро (1876) обнаружил на спинномозговой лягушке, что раздражение кожи на задней лапке вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Н. А. Миславский (1887) открыл, что кора больших полушарий одновременно возбуждает нервный центр расширения зрачка и тормозит тонус нервного центра, суживающего зрачок. Ч. Шеррингтон (1894) доказал, что возбуждение центров III и IV черепно-мозговых нервов тормозит центр VI нерва. Он назвал этот факт реципрокной иннервацией. В школе Н. Е. Введенского реципрокная иннервация обозначается как сопряженная.
Н. Е. Введенский (1896) установил, каким образом осуществляется взаимодействие двух очагов возбуждения в моторных нервных центрах коры больших полушарий головного мозга. Если раздражать на одном полушарии точку для сгибателей, то возбуждение нервного центра сгибателей на раздражаемой стороне вызывает на противоположной стороне сопряженное торможение нервного центра сгибателей и возбуждение нервного центра разгибателей.
Взаимодействие возбуждения и торможения — общее свойство всей нервной системы — обнаруживается и на спинном мозге. В зависимости от частоты и силы раздражения рецепторов и лабильности нервных центров в них в каждый момент получается возбуждение или торможение, а в соседних центрах в это время возникает торможение или возбуждение, что обеспечивает рефлекторную координацию движений. В результате взаимного сочетания возбуждения и торможения в соседних нервных центрах одни и те же мышцы в каждый момент могут быть антагонистами или синергистами. Один и тот же моторный нейрон при различных условиях раздражения и лабильности может находиться или в состоянии возбуждения или в состоянии торможения.
Закон иррадиации возбуждения в сторону очага повышенной возбудимости и принцип доминанты А. А. Ухтомского
Факты реципрокных (сопряженных) отношений между нервными центрами, именно торможение возбужденным центром других нервных центров, были установлены в прошлом веке. В. М. Бехтерев (1903) обнаружил, что импульсы, поступающие по афферентным волокнам, сосредоточиваются в определенных, центрах, что приводит к торможению других центров — рефлекс сосредоточения. И. П. Павлов (1903) утверждал, что сильное возбуждение одного центра отвлекает возбуждение от других центров, что приводит к образованию временной нервной связи, соответствующей акту внимания. Волны, возбуждения оптимальной силы, поступающие в нервный центр в оптимальном ритме, создают в нем очаг повышенной возбудимости. Возбудимость нервного центра повышается также гуморальным путем — действием гормонов и других веществ. Возбуждение иррадиирует в сторону очага повышенной возбудимости, и таким образом создается «господствующий очаг возбуждения».
Доминантой А. А. Ухтомский (1923) назвал «временно господствующий рефлекс, которым трансформируется и направляется для данного времени, при прочих равных условиях, работа прочих рефлекторных дуг и рефлекторного аппарата в целом». Принцип доминанты является «общим рабочим принципом нервных центров».
Господствующий очаг возбуждения вызывает в соседних нервных центрах торможение вследствие одновременной индукции. Чем больше возбуждение в очаге доминанты, тем больше заторможены соседние нервные центры. При осуществлении двух синергических рефлексов, имеющих общий конечный путь, рефлекс усиливается. При раздражении двух различных рецептивных полей вызываются два разных рефлекса, из которых один может вызвать торможение другого. В этом случае в нервной системе одновременно возникают два очага возбуждения, один из которых доминирует над другим.
По теории доминанты, главное значение имеет не чисто анатомический принцип общего конечного пути (Ч. Шеррингтон), а динамика одновременных или последовательных возбуждений в нервных центрах. Чем выше возбудимость и чем больше стойкость возбуждения очага доминанты, тем больше его способность суммировать приходящие к нему импульсы и тем сильнее возбуждение. Каждое слабое подпороговое раздражение создает в очаге доминанты повышение возбудимости и тем самым изменяет результат действия последующих посторонних импульсов. Посторонние для доминирующего очага раздражители, вызывающие другие рефлексы, не только не мешают доминанте, но, наоборот, подкрепляют, усиливают доминанту и тем самым увеличивают торможение в других сопряженных рефлекторных кольцах.
Следовательно, очаг доминанты характеризуется: 1) повышенной возбудимостью; 2) способностью к суммированию и накоплению возбуждения при притоке к нему импульсов, вызывающих другие рефлексы; 3) по мере развития в нем возбуждения способностью тормозить текущие рефлексы, встречаемые на общем конечном пути; 4) стойкостью возбуждения. Длительное возбуждение очага доминанты поддерживается слабыми раздражителями и не исчезает сразу после прекращения их действия (инерция доминанты). Доминанта обладает значительной инерцией, пока не будет выполнен соответствующий доминирующий рефлекс. Когда доминирующий рефлекс осуществляется длительно, то его инерция преодолевается лишь активным торможением с другой достаточно устойчивой доминанты. И пятое свойство — способность к самовозобновлению.
Существует также предел возбудимости, при достижении которого даже очень слабые или редкие раздражения рецептивных полей других рефлексов могут оказаться чрезмерно сильными и частыми и вызвать переход возбуждения очага доминанты в его торможение.
Таким образом, возбуждение и торможение, развивающиеся в нервной системе, рассматриваются как единое целое, а не изолированно. Существенное значение имеет исходное физиологическое состояние нервной системы. Начавшийся рефлекс может либо усиливать, либо тормозить другие рефлексы в зависимости от условий раздражения нервной системы и ее текущей лабильности.
Наличие доминантных очагов объясняет единство и направленность движений организма, их координацию. В каждый момент совершаются определенные направленные движения, исключающие другие движения, что «достигается распределением тонуса, тетанического сокращения и торможения» (А. А. Ухтомский). Очаг доминанты может возникать в двигательной, вегетативной и воспринимающей областях. Так, жгучая боль (каузальгия) при повреждениях периферических нервов у человека, впервые описанная Н. И. Пироговым, усиливается при самых, легких прикосновениях к раненой конечности.
Теория доминанты и рефлекторная теория И. П. Павлова объясняют механизм образования условного рефлекса притягиванием возбуждения из более слабого очага при действии условного раздражителя в более сильный его очаг, который образуется при действии безусловного раздражителя.
Обе теории имеют значение и для психологии.
Теория И. П. Павлова о главенствующем в данный момент сильном очаге возбуждения в коре больших полушарий, который притягивает возбуждение из более слабых очагов возбуждения и по правилу индукции тормозит окружающие участки коры, и теория доминанты раскрывают физиологические механизмы психического акта внимания. Смена одной доминанты другой проявляется в переключении внимания. Психологические исследования подтверждают теорию доминанты и показывают, что разнообразные слабые раздражения усиливают концентрацию внимания и что умственная работа протекает в обычной обстановке лучше, чем в абсолютной тишине. Для педагогов очень важно то, что посторонние раздражения небольшой силы, нарушающие монотонность, увеличивают общий тонус организма и способствуют концентрации внимания.
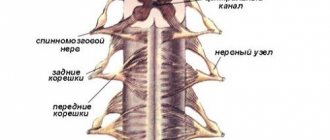
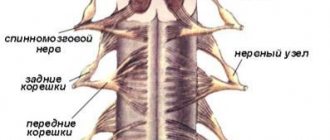
Спинномозговые корешки
Связь спинного мозга с периферией осуществляется посредством нервных волокон, проходящих в спинномозговых корешках, по ним поступают к спинному мозгу афферентные импульсы и проходят от него на периферию эфферентные импульсы. По обеим сторонам спинного мозга имеется по 31 паре передних и задних корешков.
Функции спинномозговых корешков были выяснены при помощи методов перерезки и раздражения и подтверждены путем отведения электрических потенциалов. Передние спинномозговые корешки содержат центробежные, эфферентные волокна, а задние — центростремительные, афферентные волокна. Этот факт получил название закона распределения афферентных и -эфферентных волокон в спинномозговых корешках, или закона Мажанди (по имени физиолога, впервые описавшего соответствующие наблюдения).
После односторонней перерезки всех передних корешков у лягушки или у любого другого животного исчезают рефлекторные движения соответствующей половины тела, чувствительность же ее сохраняется. Перерезка задних корешков не влечет за собой утраты способности к движению, но чувствительность в участках тела, которые снабжались соответствующими корешками, исчезает.
Эффектное доказательство функциональной роли передних и задних корешков было дано И. Мюллером, который перерезал у лягушки с одной стороны спинного мозга передние, а с другой стороны — задние корешки, иннервирующие задние конечности. На той стороне тела, где были перерезаны передние спинномозговые корешки, лапка висела как опущенная плеть, однако раздражение ее вызывало движении других частей в частности противоположной конечности. На другой же стороне, где были перерезаны задние корешки, лапка производила движение в ответ на раздражение других участков тела, но не реагировала на раздражение ее самой вследствие полной потери чувствительности.
В дальнейшем было показано, что в передних корешках прохо кроме моторных нервов скелетной мускулатуры, также другие эффере ные нервные волокна: сосудистые и секреторные, а также идущие к г коп мускулатуре. Их наличие не противоречит закону Мажанди, так все они являются эфферентными.
Парадоксальным, на первый взгляд, является тот факт, что раздражение передних корешков часто сопровождается ощущением боли. Однако этот факт не противоречит закону Мажанди, так как установлено, что часть волокон, проходящих в за корешках, заворачивает в передние и направляется в спинномозговые оболочки, снабжая их чувствительными окончаниями. В спинной мозг они вступают так же, как и остальные афферентные нервы, через задние корешки. В этом можно убедиться, перерезав несколько задних корешков и раздражая после этого соответствующие им передние корешки: болевых ощущений, которые получили название возвратной чувствительности, при этом уже не наблюдается.
Волокна, входящие в состав передних корешков, представляют собой аксоны моторных клеток передних рогов, а также относящихся к вегетативной нервной системе клеток, расположенных в боковых рогах рогах грудных и поясничных сегментов спинного мозга. Волокна, образующие задние корешки, представляют собой отростки биполярных клеток межипозвоночных специальных ганглиев.
Месторасположение тел нейронов, от которых берут начало волокна, проходящие в спинномозговых корешках, устанавливается посредством опытов, в которых производится перерезка корешков или ограниченное повреждение серого вещества снинного мозга, а затем через несколько дней на гистологических срезах прослеживается перерождение нервных волокон.
Перерезка заднего корешка ниже спинномозгового узла влечет за собой перерождение идущих на периферию волокон, перерезка же выше узла имеет следствием перерождение волокон, входящих в спинной мозг. В области самого узла нервные волокна не перерождаются, что свидетельствует о том, что здесь находятся тела нервных клеток, от которых берут начало волокна задних корешков. Волокна же передних корешков перерождаются к периферии от места перерезки при любом ее уровне, а также при повреждении передних или боковых рогов. Это показывает, что в последних расположены тела нейронов, отростки которых проходят в составе передних корешков.
В спинномозговых корешках находятся нервные волокна разной толщины, имеющие разную скорость проведения.
В задних корешках находятся толстые волокна, относящиеся к группе Аα, которые являются афферентными проводниками, идущими от ядерной сумки мышечных веретен и телец Гольджи, расположенных в сухожилиях. Импульсы, проходящие по этим волокнам, вызывают миотатические рефлексы, возникающие в ответ на растяжение мышцы. Волокна средней толщины (5—12 мк), относящиеся к типу Аβ и Аγ, проходящие по задним корешкам. идут от тактильных рецепторов и от рецепторов мышечных веретен, расположенных к периферии от ядерной сумки. Подобные же волокна идут от рецепторов полых внутренних органов (мочевого пузыря, желудка, тонких и толстых кишок, прямой кишки и др.).
Афферентные волокна Аβи Аγ, несут импульсы от мехагорецепторов. После вступления в спинной мозг эти волокна заходят в задние столбы, отдавая коллатерали вставочным (так называемым комиссуральным) нейронам, расположенным в сером веществе выше- и нижележащих сегментов спинного мозга. Импульсы, поступающие по небольшому числу афферентных волокон этой группы, способны вызвать возбуждение большого числа нейронов спинного мозга. Тем самым раздражение ограниченного количества рецепторов, например при уколе пальца, может вызвать сокращение большой группы мыши, приводящее к сгибанию руки или ноги. Наиболее тонкие волокна (диаметром 2—5 мк) задних корешков, относящиеся к группе АΔ , несут импульсы от терморецепторов и болевых рецепторов. Импульсы от последних поступают к спинному мозгу также по тонким безмиелиновым волокнам, принадлежащим к группе С.
В передних корешках проходят эфферентные нервные полокна также разных типов. Здесь имеются:
- толстые волокна (в среднем диаметром 16 мк) тина Аα, несущие импульсы к скелетной мускулатуре;
- тонкие волокна (в среднем диаметром 8 мк) типа Аγ, иннервирующие сократительные элементы мышечного веретена,
- преганглионарные симпатические волокна, относящиеся к типу В.
После перерезки задних корешков наряду с исчезновением чувствительности наблюдаются также расстройства движения. Так, если перерезать с обеих сторон спинного мозга все задние корешки, иннервирующие задние конечности собаки, сохранив в целости передние корешки, то животное в первое время после операции теряет способность передвигаться при помощи этих конечностей. Через некоторое время движения задних лап, потерявших чувствительность, восстанавливаются, но имеют ненормальный характер: движения порывистые, резкие; лапы чрезмерно сильно сгибаются и также сильно разгибаются. Такие движения называются атактическими. Они встречаются и у человека при заболеваниях спинного мозга, сопровождающихся поражением восходящих путей (спинномозговая атаксия).
Расстройство координации движений наступает вследствие прекращения потока афферентных импульсов в мозг, прежде всего от рецепторов двигательного аппарата, т. е. от проприорецепторов, а также от экстерорецепторов кожи. Отсутствие информации о состоянии двигательного аппарата в каждый данный момент движения приводит к тому, что мозг теряет способность контролировать, оценивать характер движения и вносить поправки на всех этапах двигательного акта. И хотя эфферентные импульсы поступают из мозга в мышцы и вызывают их сокращения, процесс этот не контролируется и не регулируется, так как отсутствует обратная связь, без которой невозможно управление двигательными актами и выполнение точных и плавных движений. Вот почему нарушаются двигательные акты, требующие точных движений рук, например игра на пианино или письмо, после анестезии, т. е. понижения или исчезновения чувствительности кожи руки от холода или после внутрикожного введения кокаина-яда, парализующего рецепторы. Потеря чувствительности приводит, кроме того, к ослаблению мышечного тонуса.