Classification of nerve fibers
Classification of nerve fibers by diameter and conduction speed
− classification of motor and sensory nerve fibers according to Erlanger-Gasser;
− classification of sensory nerve fibers according to Lloyd-Hunt;
− compare Erlanger classifications of nerve fibers
− Gasser and Lloyd-Hunt with the classification of nerve fibers according to De Meyer (Tables 1, 2, 3).
Types of nerve fibers and their functions
When recording the electrical activity of the nerve trunk, Joseph Erlanger and Herbert Gasser in 1937 discovered the composite nature of the current action of the mixed nerve.
Based on the data obtained (diameter, conduction velocity, function), a classification of nerve fibers was developed (Table 1, summary Table 3), according to which all nerve fibers are divided into groups A, B and C (Latin letters) with further gradations (α, β, etc.).
Erlanger-Gasser classification of nerve fibers - classification of motor and sensory nerve fibers:
A, B, C - groups of fibers;
α, β, γ, δ - subgroups of group A fibers.
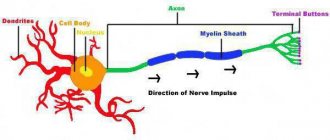
Type A fibers are thick myelinated nerve fibers.
The thickest of them, Aα, have a diameter of 12–22 μm and an excitation speed of 70–120 m/s. These fibers conduct excitation from the spinal motor nerve centers (motor centers of the spinal cord) to the skeletal muscles (motor fibers) and from muscle receptors to the corresponding nerve centers.
Other groups of type A fibers (β, γ, δ) have a smaller diameter - from 8 to
1 µm and lower excitation speed - from 5 to 70 m/s. The fibers of these groups predominantly conduct excitation from various receptors (tactile, temperature, pain; receptors of internal organs or visceroreceptors) in the central nervous system, with the exception of γ-fibers, a significant part of which conduct excitation from the spinal cord to intrafusal muscle fibers.
Table 1
Classification of nerve fibers according to Erlanger-Gasser
| Type fibers | Function (optional) | Average diameter, µm | Average conduction speed, m/s |
| Aα | Primary afferents of muscle spindles, motor fibers of skeletal muscles | 15 | 100 (70 – 120) |
| Aβ | Cutaneous afferents of touch and pressure | 8 | 50 (30 – 70) |
| Aγ | Motor fibers of muscle spindles | 5 | 20 (15 – 30) |
| Aδ | Cutaneous afferents of temperature and pain | <3 | 15 (12 – 30) |
| IN | Sympathetic preganglionic fibers | 3 | 7 (3 – 15) |
| WITH | Cutaneous pain afferents Sympathetic postganglionic fibers | 1 (unmyelinated) | 1 (0,5 – 2) |
Type B fibers are myelinated preganglionic fibers of the autonomic nervous system. Their diameter is 1–3.5 microns, and the excitation speed is 3–18 m/s.
Type C fibers include unmyelinated nerve fibers of small diameter - 0.5 - 2 microns. The excitation speed in these fibers is no more than 3 m/s (0.5 - 3 m/s).
Most type C fibers are postganglionic fibers of the sympathetic division of the autonomic nervous system, as well as nerve fibers that conduct excitation from pain receptors, some thermoreceptors, and pressure receptors.
Lloyd-Hunt classification of nerve fibers
The Lloyd-Hunt classification is widely used for sensory nerve fibers (Roman numerals), which is presented in Table 2 and Summary Table 3.
table 2
Classification of nerve fibers according to Lloyd-Hunt
Fiber type | Function (optional) | Average diameter, µm | Average conduction speed, m/s |
| I | Primary afferents from muscle spindles and tendon organs | 13 | 75 (70 – 120) |
| II | Cutaneous mechanoreceptors | 9 | 55 (25 – 70) |
| III | Muscle deep pressure sensors | 3 | 11 (10 – 25) |
| IV | Unmyelinated pain afferents | <3 |
Physiology of neuromuscular tissue
It is known that under the influence of a stimulus, living cells and tissues move from a state of physiological rest to a state of activity. The greatest response among tissues to irritation is observed from nervous and muscle tissue. The main properties of nervous and muscle tissue are excitability, excitation, conductivity, refractoriness and contractility.
Excitability is the ability of living tissue to respond to the action of a stimulus by changing physiological properties and the occurrence of the excitation process. Excitation is an active physiological process that occurs in tissue under the influence of stimuli and is characterized by changes in the level of metabolic processes in tissues, release of energy, contraction of muscle tissue, secretion, and generation of a nerve impulse.
Conductivity is the ability of living tissue to conduct excitation waves (biopotentials). Refractoriness is a temporary decrease in tissue excitability that occurs as a result of excitation. Lability is the dependence of tissue on the characteristics of metabolic processes, which can be excited a certain number of times per unit of time.
There are electrical, chemical, mechanical and thermal stimuli that can cause a response from excitable tissues. According to biological characteristics, stimuli can be adequate and inadequate, and according to strength - subthreshold, threshold and suprathreshold.
The strength of the stimulus (the law of irritation) is of decisive importance for the appearance of excitation. There is a certain relationship between the strength of irritation and the response. The greater the strength of the stimulus, the higher, to the appropriate level, the response from the excitable tissue.
The duration of the stimulus is also of great importance. The relationship between the strength of the stimulus and the duration of its action required for the appearance of a minimal response is determined by the force-time curve. The minimum current (voltage) that can cause excitation is called rheobase (ordinate segment (OA). In addition to rheobase, an important parameter of the force curve is chronoxia.
The latter reflects the shortest period of time during which a current equal in strength to twice the rheobase (abscissa segment OF) causes excitation in the tissue. The magnitude of chronoxia is used to judge the speed at which excitation appears in the tissue: the lower the chronoxia, the faster excitation occurs.
The adaptation of excitable tissue to a slow increase in the strength of the stimulus is called accommodation. The latter is due to the fact that as the strength of the stimulus increases, active changes occur in the tissue, which increase the threshold of irritation and prevent the development of excitation. Thus, the rate of increase in irritation over time is called the irritation gradient.
The law of the gradient of irritation is a reaction to a stimulus, which depends on the urgency or steepness of the increase in the stimulus over a certain time: the higher the gradient of irritation, the stronger (up to certain limits) the response of the excitable object.
The resting potential (membrane potential) is the potential difference between the outer surface of the cell and its internal contents; it is about 60-90 mV, depending on the characteristics of a particular cell.
An action potential (excitation potential) occurs when an area of a nerve or muscle fiber is exposed to a stimulus of sufficient strength and duration. A manifestation of the resulting excitation is a rapid fluctuation of the membrane potential. In this case, the excited area has an electronegative charge relative to the non-excited one.
In the action potential, local oscillations of the membrane potential, the peak of the action potential and trace potentials - negative and positive - are distinguished. The peak of the action potential is a short-term change in the internal potential and has a very fast rising phase and a somewhat slow falling phase.
Following the peak of the action potential, weaker and longer negative and then positive trace potentials are recorded. The duration of the action potential in nerve and muscle fibers is 0.1-5.0 ms. A system of penetrating channels for Na+, K+, C1-, Ca2+ ions plays a role in the development of the resting potential and action potential. The nerve membrane contains specific sodium, potassium, chloride and calcium channels that allow only the above ions to pass through.
These channels have gate mechanisms and can be open or closed. Determining the state of membrane ion channels is very important for the formation of the resting potential, where the leading role is played by the uneven distribution of potassium ions. Sodium ions play a major role in the formation of an action potential. Conducting excitation is a specialized function of nerve fibers.
The rate of excitation along them depends mainly on the diameter and histological features of the structure of the nerve fibers. The larger the diameter of the nerve fiber, the higher the speed of propagation of excitation in it. For example, along a nerve fiber with a diameter of 12-22 microns, the speed of propagation of excitation is 70-120 m/s, and along a nerve fiber with a diameter of 8-12 microns - only 40-70 m/s.
Based on their histological structure, nerve fibers are divided into myelinated and non-myelinated. The myelin fiber consists of an axial cylinder and the myelin, or Schwann, sheath covering it. It contains fatty substances that have high resistivity and play an insulating role. At regular intervals, the myelin sheath is interrupted and leaves open areas of the axial cylinder about 1 μm wide (the node of Ranvier).
The surface of the axial cylinder is represented by the plasma membrane, and its contents are axoplasm. Unmyelinated fibers do not have a myelin sheath, but are covered only by Schwann cells. The space between the Schwann cells and the axial cylinder is filled with intercellular fluid, which allows the surface membrane of the axial cylinder to communicate with the environment surrounding the nerve fiber.
The transmission of excitation along myelinated and non-myelinated fibers has its own characteristics. Thus, the transmission of the action potential along the myelin fiber occurs spasmodically from one node of Ranvier to another, which allows excitation to spread without extinction.
The speed of propagation along myelinated fibers is much higher than through non-myelinated ones. If the excitation speed along motor nerve fibers (covered with a myelin sheath) is 80-120 m/s, then along fibers that do not have myelin fibers it is only 0.5-2.0 m/s. The propagation of excitation along the nerve fiber when excitation is applied obeys the corresponding laws.
The law of physiological integrity - the conduction of excitation along a nerve fiber is possible only if not only its anatomical, but also physiological integrity (continuity) is preserved.
The law of two-way conduction of excitation - the transfer of excitation occurs in two directions - centripetal and centrifugal.
The law of isolated conduction of excitation - when stimulation is applied, excitation is carried out only along one nerve fiber and does not cover adjacent fibers, which determines strict coordination of reflex activity. Nerve fibers get tired a little. This is due to low energy costs and fast recovery processes.
A synapse is a specialized structure that ensures the transmission of a nerve impulse from a nerve fiber to an effector cell - a muscle fiber, neuron or secretory cell.
Synapses are classified according to anatomical and histological principles (neuro-secretory, neuromuscular, interneuronal); neurochemical principle (adrenergic - the mediator norepinephrine and cholinergic - the mediator acetylcholine); functional (excitatory and inhibitory). The neuromuscular junction consists of three main structures: the presynaptic membrane, the synaptic cleft, and the postsynaptic membrane.
The presynaptic membrane covers the nerve ending, and the postsynaptic membrane covers the effector cell. Between them there is a synaptic cleft. The postsynaptic membrane differs from the presynaptic membrane in that it has protein chemoreceptors that are sensitive not only to mediators and hormones, but also to drugs and toxic substances. The structure of the neuromuscular synapse determines its physiological properties:
- unilateral conduction of excitation (from pre- to postsynaptic membrane) in the presence of transmitter-sensitive receptors only in the postsynaptic membrane;
- synaptic delay in the conduction of excitation associated with the low rate of diffusion of the transmitter in comparison with the speed of the nerve impulse;
- low lability and high fatigue of the synapse;
- high selective sensitivity of the synapse to chemicals.
The transmission of excitation at the synapse is a complex physiological process that goes through several stages:
- mediator synthesis;
- mediator secretion;
- interaction of the mediator with receptors of the postsynaptic membrane;
- inactivation (complete loss of activity) of the mediator.
It is known that some chemicals, including drugs, can significantly affect excitation at the synapse. This phenomenon has found application in clinical practice.
A neuromotor unit is an anatomical and functional unit of skeletal muscle, which consists of an axon (the long extension of a spinal cord motor neuron) and a certain number of muscle fibers innervated by it. A neuromotor unit may contain a different number of muscle fibers (from a few to several thousand), which depends on the specialization of the muscle. The motor unit works as a single unit. The impulses generated by the motor neuron activate all the muscle fibers that form it.
The main function of skeletal muscles is contraction, which is expressed by various human movements. Skeletal muscles also perform receptor, metabolic and thermoregulatory functions. They are formed by a large number of multinucleated muscle fibers.
The contractile part of the muscle fiber are long muscle filaments - myofibrils, which run inside the fiber from one end to the other and have a transverse outline. The latter is formed by alternating dark (anisotropic) A-disks and light (isotropic) 1-disks. A Z-line passes through the middle of the 1-disk; two adjacent Z-lines delimit the sarcomere, a structural and functional unit. Electron microscopy of the fibers shows that the A-disk contains a lighter area (H-zone), and in the center this disk is crossed by a dark stripe—the M-line.
The dark disk is formed by thick filaments of the myosin protein, and the light 1-disc is formed by thin filaments of the actin protein. Muscle fiber also contains fibrillar rod-shaped protein - tropomyosin and globular protein - troponin. The contraction mechanism consists of moving (pulling) thin filaments along thick filaments towards the center of the sarcomere due to transverse actinomyosin bridges. The main source of energy required for muscle contraction is adenosine triphosphoric acid (ATP) and the presence of Ca2+ and Mg2+ ions.
The conversion of chemical energy into mechanical energy occurs in the muscle both without oxygen and with its participation. The anaerobic (oxygen-free) phase is characterized by a series of sequential reactions that lead to the breakdown of ATP and creatine phosphate, and their restoration. The energy released in this case is used for muscle contraction and restoration (resynthesis) of these substances. The aerobic (oxygen) phase of chemical transformations is associated with the oxidation of lactic acid to carbon dioxide and water. The resulting energy is used to further convert lactic acid residues into glucose and then into glycogen.
The activity of skeletal muscles is regulated by the central nervous system - the cerebral cortex, through sensory, motor and sympathetic nerve fibers. Skeletal muscles have the following physiological properties: excitability, conductivity, refractoriness, lability and contraction. Excitability, the speed of spread of excitation, and the lability of muscle tissue are lower than in nervous tissue, and the refractory period is longer than the nervous one.
Skeletal muscles can perform work in isotonic, isometric and auxotonic contraction modes. During the first contraction, the muscle fiber is mainly shortened, but the tension remains constant, and during the second contraction, the length of the muscle fibers remains the same, but the length and tension change. The nature of skeletal muscle contraction depends on the frequency of stimulation (the frequency of nerve impulses).
Irritation by a single impulse leads to a single muscle contraction, and nerve impulses following one after another lead to tetanic contraction, or tetanus.
The physiological properties of smooth muscles are associated with the peculiarities of their structure, the level of metabolic processes and differ significantly from the characteristics of skeletal muscles. Smooth muscles are less excitable than striated muscles. Smooth muscle contraction occurs more slowly and lasts longer. The refractory period of smooth muscles is longer than that of skeletal muscles (up to several seconds).
A characteristic feature of smooth muscles is their ability to perform automatic activity, which is provided by nerve elements. Smooth muscles are innervated by sympathetic and parasympathetic autonomic nerves and are highly sensitive to some biologically active substances (acetylcholine, adrenaline, norepinephrine, serotonin, etc.).
Myelination
The nerve fiber is based on the process of a neuron, which forms a kind of axis. Outside, it is surrounded by a myelin sheath with a biomolecular lipid base consisting of a large number of turns of mesaxon, which spirals onto the neuronal axis. Thus, myelination of nerve fibers occurs.
The myelinated nerve fibers of the peripheral system are additionally covered on top with auxiliary Schwann cells that support the axon and nourish the body of the neuron. The surface of the pulp membrane has intervals - nodes of Ranvier, in these places the axial cylinder is attached to the outer Schwann membrane.
The myelin layer does not have electrically conductive properties; the interceptions have them. Excitation occurs in the interval of Ranvier closest to the place of influence of the external stimulus. The impulse is transmitted spasmodically, from one interception to another, this ensures a high speed of impulse propagation.
Myelinated nerve fibers regulate metabolism in muscle tissue and have high resistance to bioelectric current.
The gaps of Ranvier generate and amplify impulses. The fibers of the central nervous system do not have a Schwann membrane; this function is performed by oligodendroglia.
The pulpless tissues have several axial cylinders, they do not have a myelin layer and interceptions, they are covered with Schwann cells on top, and slit-like spaces are formed between them and the cylinders. The fibers have poor insulation, allow the propagation of an impulse from one neuron process to another, are in contact with the environment throughout, the speed of impulse conduction is much lower than that of pulpy fibers, and the body requires more energy.
Large nerve trunks are formed from the pulpal and non-pulpate processes of neurons, which, in turn, branch into smaller bundles and end with nerve endings (receptor, motor, synapses).
Nerve endings are the ends of myelinated and unmyelinated nerve fibers that form interneuron contacts, receptor and motor endings.
Laws of excitation conduction in nerve fibers and nerves
Classification of nerve fibers according to Erlanger-Gasser
In 1939, American [Mf54] physiologists Joseph Erlanger [B55] and Herbert S. Gasser [B56] [B57] recorded action currents from the entire nerve trunk of the frog sciatic nerve at different distances from the stimulating electrode (Fig. 210041905).[B58]
It was found that the recorded total potential has a number of peaks, which were designated by the letters of the Latin alphabet A, B, C (Fig. , I). Peak A had additional peaks labeled with the Greek letters α , β , γ , δ (Fig. ,II). [B59] In 1944, the work of J. Erlanger and G.S. Gasser was awarded the Nobel Prize [B60].
Nerves in vertebrates consist of three main groups of fibers
(A, B and C), differing in the degree of myelization, fiber diameter, duration of the AP peak (AP development rate), electrical excitability, its compensation and conduction speed (all these indicators in the series A - B - C fall).
Group A
includes the thickest well-myelinated motor and sensory fibers;
group B -
weakly myelinated, preganglionic fibers of the autonomic nervous system;
group C -
unmyelinated, postganglionic (sympathetic) fibers.
In group A, in the row a, b, g, d, these indicators also fall. The ratios of the properties of these groups of fibers are shown in table. .
It should be noted that the indicated ratios of the thresholds of electrical stimulation of groups of fibers do not reflect the exact ratio of the electrical excitability of their membranes. The relatively high thresholds of thin fibers when they are irritated in the nerve trunk are determined mainly by the fact that thin fibers have a higher input resistance compared to thick ones. They contain such a small part of the irritating current that at the threshold strength for Aa
-fibers, it is completely insufficient to create any significant depolarization on the membrane of thinner fibers.
For the same reason (high RI
), the PD of thin fibers removed from the trunk (extracellularly) appears significantly smaller than the PD of thick fibers.
Rice. . Components of the mixed nerve action potential.
I – at a relatively slow recording speed.
II – at a relatively high recording speed.
Explanation in the text. The abscissa axis is time, the ordinate axis is the amplitude of the compound potential in mV.
Total electrical activity of the nerve
is created by its fibers, each of which generates its own PD, standard in amplitude and time parameters, spreading in both directions from the point to which the stimulation is applied. The total electrical signal of the nerve depends on the number of active fibers, the synchrony of their activity, the method of abduction and other circumstances.
Let us consider the case of the so-called single-phase derivation,
in which one output electrode (active) is located on a normal section of the trunk, and the other (indifferent) is located on a damaged one, where the fibers are completely depolarized (Fig. 1.19).
Let artificial stimulation be used and the stimulating electrode (cathode) be located sufficiently close (l =
3 mm) to the active output electrode, and the nerve is placed in a non-conducting medium (oil or air). Here, a strong single stimulation leads to synchronous excitation of all fibers, while the active output electrode records the total action potential of the nerve, which is similar in shape to the action potential of an individual A (alpha) fiber, but is slightly more prolonged in time. This nerve action, however, does not follow the all-or-none rule. With threshold stimulation it is negligibly small, with increasing stimulus strength it gradually increases, reaching a maximum equal to 5-10 mV under normal conditions, and 50-100 mV under sucrose bridge conditions.
With a further increase in stimulus strength, this AP slightly lengthens in time. All changes in the amplitude and duration of the AP nerve peak with increasing stimulus are determined by an increase in the number of active fibers, the connection to low-threshold and fast A (alpha) fibers of higher threshold slow beta, gamma, delta fibers of group A, then B and, finally, C-groups.
| Fiber groups (according to Erlanger and Gasser) | Diameter, microns | Conduction speed, m/s |
| Aa | 13 — 22 | 70 — 120 |
| Ab | 8 -13 | 40 — 70 |
| Ag | 4 — 8 | 15 – 40 |
| Ad | 1 – 4 | 5 – 15 |
| B | 1 — 3 | 3 – 14 |
| C | 0,5 – 1,0 | 0,5 — 2 |
Table . Classification of nerve fibers according to J. Erlanger and H. Gasser
| Fiber groups (according to Erlanger and Gasser) | Diameter, microns | Electrical stimulation thresholds (relative to Aa) | PD1 peak duration | Negative Trace Potential (NTP) | Positive trace potential [B61] | Conduction speed, m/s | |
| Duration, ms | SP amplitude, % to AP amplitude | Duration, ms | SP amplitude, % to AP amplitude | ||||
| Aa | 13 — 22 | 1,0 | 0,4 | 15 – 20 | 40 — 60 | 0,2 | 70 — 120 |
| Ab | 8 -13 | 40 — 70 | |||||
| Ag | 4 — 8 | 15 – 40 | |||||
| Ad | 1 – 4 | 5 – 15 | |||||
| B | 1 — 3 | 11,7 | 1,2 | no OSB | 100 – 300 | 3 – 14 | |
| C | 0,5 – 1,0 | 100,0 | 2,0 | 50 – 60 | 300 — 1000 | 0,5 — 2 |
1Absolute refractory phases have approximately the same value
The basic properties of autowaves regarding their propagation also apply to the action potentials of nerve fibers:
1. propagates without attenuation both along the length of the fiber and when it branches (Fig. 709170042).
2. do not interfere (Fig. 709170043).
3. do not reflect from obstacles (Fig. 709170044).
4. The direction of propagation is determined by the zones of refractoriness and rest, ensuring bilateral conduction of excitation (Fig. 709170045, 709170046).
Rice. 709170042. Distribution of APs during branching of nerve fibers. The correct option and the wrong option, which is often found in student answers, are shown.
Rice. 709170043. Distribution of PD at the convergence of nerve fibers. The correct option and the wrong option, which is often found in student answers, are shown.
Rice. 709170044. PDs are not reflected from obstacles. The correct option and the wrong option, which is often found in student answers, are shown.
The AP travels equal distances (L) from the location of the stimulus in the same time (t) and maintains its amplitude, other things being equal.
Rice. 709170045. Bilateral conduction along nerve fibers. L
– distance from the place of stimulus action,
t
– time of AP conduction from the place of stimulus action to the location of the recording electrodes,
A
– AP amplitude.
The condition for maintaining the anatomical and physiological continuity of the fiber is usually emphasized
Anatomical and physiological fiber continuity
Conduction of impulses is possible only if the fiber is anatomically intact, therefore both cutting of nerve fibers and any injury to the surface membrane disrupt conduction. Nonconductivity is also observed when the physiological integrity of the fiber is disrupted (blockade of sodium channels of the excitable membrane by tetrodotoxin or local anesthetics, sudden cooling, etc.). Conduction is also disrupted during persistent depolarization of the nerve fiber membrane by K+, which accumulates in the intercellular spaces during ischemia. Mechanical trauma, compression of the nerve during inflammatory tissue edema may be accompanied by partial or complete disruption of conduction function.