The grooves and convolutions of the medial and inferior surfaces of the hemisphere. Localization of nerve centers.
Medial surface of the cerebral hemisphere
formed by all its lobes except the insular lobe.
The groove of the corpus callosum (sulcus corporis callosi)
goes around it from above, separating the corpus callosum from
the lumbar gyrus (gyrus cinguli).
Then this groove is directed downward and forward and continues into
the hippocampal groove (sulcus hippocampi).
groove (sulcus cinguli)
runs above which begins in front and below the beak of the corpus callosum.
As it rises, the groove turns posteriorly and runs parallel to the groove of the corpus callosum. At the level of its ridge, its marginal branch (ramus marginalis) extends upward from the cingulate sulcus,
and the sulcus itself continues into
the subparietal sulcus (sulcus subparietialis).
Posteriorly and inferiorly from the splenium of the corpus callosum, the cingulate gyrus narrows and forms
the isthmus gyri cinguli.
The marginal branch of the cingulate sulcus limits
the pericentral lobule (lobulus paracentralis) behind,
and
the precuneus (preciuneus) in front,
which belongs to the parietal lobe.
Down and posteriorly through the isthmus, the cingulate gyrus passes into the parahippocampal gyrus (gyrus parahippocampalis),
which ends in front
with a hook (uncus)
and is bounded above
by the hippocampal sulcus
(
sulcus hippocampi).
The cingulate gyrus, isthmus and
parahippocampal gyrus
are combined under the name
vaulted gyrus (gyrus fornicatus).
The dentate gyrus (gyrus dentatus)
is located deep in the hippocampal sulcus On the medial surface of the occipital lobe, two deep grooves are located at an acute angle to each other:
the parieto-occipital groove (sulcus parietooccipitalis),
separating the dark lobe from the occipital lobe, and
the calcarine groove (sulcus calcarinus).
The latter begins on the medial surface of the occipital pole and goes forward to the isthmus of the cingulate gyrus.
The area of the occipital lobe lying between the parieto-occipital and calcarine grooves and having the shape of a triangle with its apex facing the junction of these grooves is called a wedge (cuneus).
The calcarine groove, clearly visible on the medial surface of the hemisphere, limits
the lingual gyrus (gyrus lingualis) above,
extending from the occipital pole posteriorly to the lower part of the isthmus of the cingulate gyrus.
Below the lingual gyrus there is a collateral groove (sulcus collateralis),
which already belongs to the lower surface of the hemisphere.
The lower surface of the cerebral hemisphere
has the most complex relief.
In front is the lower surface of the frontal lobe, behind it is the temporal pole and the lower surface of the temporal and occipital lobes, between which there is no clear boundary. On the lower surface of the frontal lobe, somewhat lateral and parallel to the longitudinal fissure of the cerebrum, there is an olfactory groove (sulcus olfactorius).
Adjacent to it below are
the olfactory bulb (bulbus olfactorius)
and
the olfactory tract (tractus olfactorius),
which passes from behind into
the olfactory triangle (trigonum olfactorium),
the medial
and
lateral olfactory stripes
(
striae olfactoriae medialis
et
are visible .
Between
the longitudinal fissure of the cerebrum (fissura longitudinalis cerebri)
and
the olfactory sulcus
lobe there is
a straight gyrus (gyrus rectus).
Lateral to the olfactory sulcus lie
the orbital gyri (gyri orbitales).
The lingual gyrus (gyrus lingualis) of the occipital lobe is limited on the lateral side by
the occipitotemporal sulcus (sulcus occipitotemporalis).
This groove passes to the inferior surface of the temporal lobe, dividing
the parahippocampal (gyrus parahippocampalis)
and
medial occipitotemporal gyrus (gyrus occipitotemporalis medialis).
Anterior to the occipitotemporal sulcus is
the nasal sulcus (sulcus rhinalis),
which bounds the anterior end of the parahippocampal gyrus -
the uncus.
The occipitotemporal sulcus separates
the medial
and
lateral occipitotemporal gyri (gyri occipitotemporalis medialis
et
lateralis).
On the medial and lower surfaces of the cerebrum there are a number of formations related to the
limbic system (from the Latin limbus
- border).
These are the olfactory bulb, olfactory tract, olfactory triangle, anterior perforated substance, mammillary bodies
located on the lower surface of the frontal lobe (peripheral part of the olfactory brain), as well as
the cingulate, parahippocampal
(together with
the hook)
and
dentate gyri
.
The subcortical structures of the limbic system are the amygdala, septal nuclei
and
anterior thalamic nucleus.
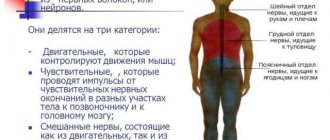
Examination card No. 72 1. The structure of a typical vertebra. Differences between the vertebrae of different parts of the spinal column. Variability of the transitional parts of the spinal column.2. Trachea and main bronchi, their topography, structure of the walls. Age characteristics and anomalies. Blood supply, venous outflow, regional lymph nodes, innervation.3. The structure of the walls of the heart. Atrioventricular conduction system.4. Sympathetic trunk, its topography, structure, sections, connections with the spinal nerves.
1 The structure of a typical vertebra. Differences between the vertebrae of different parts of the spinal column. Variability of the transitional parts of the spinal column.
The human spine is a long, curved column made up of vertebrae lying on top of each other. The vertebral bodies are built of spongy substance, covered along the periphery with a thin compact plate. A strictly defined arrangement of the spongy crossbars according to the lines of compression and tension forces ensures the strength of the vertebrae. The strength of the spinal column as a whole depends on the connections of the vertebrae and the powerful ligamentous apparatus. The spine has an amazing combination of strength and mobility.
Regardless of their belonging to one or another part of the spine, the vertebrae are homologous, that is, they have a general structural plan, determined by the vertical position of the human body. Cervical vertebrae
usually 7,
thoracic
- 12,
lumbar
- 5,
sacral
- 5,
coccygeal
- 2-4. A newborn has 33 separate vertebrae. In an adult, the lower vertebrae fuse to form the sacrum and coccyx, so there are 28 individual vertebrae.
Vertebra _
consists of
a body (corpus vertebrae),
which faces forward and is its supporting part.
Behind the body is a vertebral arch (arcus vertebrae),
connected to the vertebral body by two
legs (pedunculi arcus vertebrae).
As a result of the connection of the body and the arch,
the vertebral foramen (foramen vertebrae) is formed.
The openings of all vertebrae, overlapping one another, form
the spinal canal (canalis vertebralis)
, in which the spinal cord is located.
The surface of the vertebral body facing the vertebral foramen is concave and has nutrient openings into which blood vessels enter. 7 processes extend from the vertebral arch, to which muscles are attached. Posteriorly along the midline is the unpaired spinous process (processus spinosus).
In the frontal plane on the right and left there is a paired
transverse process (processus transversus)
.
upper
and
lower articular processes (processus articuleres superiores
et
are directed upward and downward from the arch .
The bases of the articular processes limit
the superior
and
inferior vertebral notches (incisurae vertebrals superior et inferior).
When the vertebrae are connected to each other, the lower notch of the overlying vertebra and the upper one of the underlying one form
intervertebral foramina (foramina intervertebralia) on the right and left,
through which spinal nerves and blood vessels pass.
Cervical vertebrae (vertebrae cervicales)
human vertebrae differ from other vertebrae in their small size and a small rounded hole in each transverse process.
In the natural position of the cervical vertebrae, these holes, overlapping one another, form an intermittent canal in which, starting from the VI cervical vertebra, the vertebral artery, supplying blood to the brain, goes up. Each transverse process ends in an anterior and posterior tubercles. The anterior tubercle of the VI cervical vertebra is better developed than the tubercles of other vertebrae. Due to its proximity to the carotid artery, it received the name carotid tubercle. For bleeding in the head and neck area, the carotid artery can be pressed against it. The articular processes have a rounded, smooth surface; in the upper processes it faces backward and upward, in the lower ones it faces forward and downward. The spinous processes are short, their length increases from the II to the VII vertebrae, their ends are bifurcated, except for the VII vertebra, the spinous process of which is the longest. Its apex can be easily palpated in a living person and serves as a guide in determining the upper border of the lungs, the pleural dome. The I and II cervical vertebrae differ significantly from the rest. They articulate with the skull and bear its weight. The first cervical vertebra, or atlas ,
is devoid of a body and a spinous process (the middle part of the body, having separated from the atlas, adhered to the body of the second vertebra, forming its tooth).
The first cervical vertebra has preserved remains of the body - lateral masses (massae laterales),
the posterior
and
anterior arches of the vertebra (arcus posterior
et
arcus anterior)
extend limiting the large round vertebral foramen (Fig. 56).
Twelve thoracic vertebrae (vertebrae thoracicae)
connect to the ribs.
This leaves an imprint on their structure. On the lateral surfaces of the bodies there are located the upper
and
lower costal fossae (foveae costales superior
et
inferior)
for articulation with the heads of the ribs.
On the body of the first thoracic vertebra there is a whole fossa for articulation with the head of the second rib and half a fossa for the upper half of the head of the second rib. On the body of the II vertebra, the lower half of the fossa for the II rib and half of the fossa for the III rib, etc. are visible. Thus, the II rib and the underlying ribs up to and including the VIII rib are attached to two adjacent vertebrae. The X thoracic vertebra has only an upper semi-fossa on the body, and the XI and XII vertebrae have a whole fossa for attaching ribs, which correspond to them in number. The transverse processes of the thoracic vertebrae are long, deviated posteriorly, and their ends are thickened. On the anterior surface of each transverse process of the 10 upper thoracic vertebrae there is a costal fossa of the transverse process (fovea costalis processus transversi),
with which the tubercle of the corresponding rib articulates.
The transverse processes of the XI and XII thoracic vertebrae are short and do not have costal fossae, since they do not connect to the tubercles of the ribs. The spinous processes
of the thoracic vertebrae are longer than those of the cervical vertebrae. Their downward inclined location prevents hyperextension of the spinal column, thereby protecting the organs of the thoracic cavity from damage. The articular processes of the thoracic vertebrae are located in the frontal plane, the articular surfaces of the upper articular processes are directed backward and laterally, the lower ones are directed forward and medially.
Lumbar vertebrae. Due to the heavy load, the lumbar vertebrae (vertebrae lumbales)
have a massive body, which distinguishes them from the vertebrae of other sections.
The body of the lumbar vertebra is bean-shaped, its transverse size is larger than the anteroposterior one. The height and width gradually increase from the I to the V vertebrae. The vertebral foramen
is large, triangular in shape, with rounded corners.
The long transverse processes of the lumbar vertebrae are rudiments of ribs, fused during development with the true transverse processes. The transverse processes are located in the frontal plane, their ends are deflected posteriorly. At the junction of the rudimentary rib with the true transverse process of the lumbar vertebrae, on each side there is a small protrusion - an additional process (processus accessorius).
The spinous processes are short, flat, with thickened ends, directed backwards.
This position of the spinous processes of the lumbar vertebrae provides greater mobility of the spinal column in this area. The articular surfaces of the articular processes
are located in the sagittal plane; in the upper processes they are directed medially, in the lower ones they are directed laterally.
Each superior articular process has a small tubercle - the mastoid process (processus mamillaris).
Sacrum (os sacrum)
consists of 5 fused
sacral vertebrae (vertebrae sacrales)
.
This massive bone takes on the entire weight of the body and transfers it to the pelvic bones. The sacrum has a triangular shape. It is distinguished by a wide and thickened base of the sacrum (basis ossis sacri),
directed upward, and
the apex of the sacrum (apex o'ssis sacri),
directed downward and forward.
The anterior pelvic surface (facies pelvina)
is concave, the posterior
dorsal surface (facies dorsalis)
is convex.
The base of the sacrum has articular processes
that articulate with the lower articular processes of the fifth lumbar vertebra.
The junction of the sacrum with the body of this vertebra forms a protrusion directed forward - the promontory.
Coccyx (os coccygis)
is a homologue of the caudal skeleton of animals.
In an adult, the coccyx consists of 3-5 rudimentary coccygeal vertebrae (vertebrae coccygeae).
The tailbone is triangular in shape and curved anteriorly.
Its base
is directed upward,
the apex
downward and forward,
the body
of the coccyx articulates with the sacrum, and on its posterior surface there is a paired
coccygeal horn (cornu coccygeum).
Both horns are directed upward, towards the horns of the sacrum. In older people, the coccygeal vertebrae are fused into one bone, and in women and young people they are often connected to each other using cartilaginous plates
2 Trachea and main bronchi, their topography, structure of the walls. Age characteristics and anomalies. Blood supply, venous outflow, regional lymph nodes, innervation .
Trachea (trachea),
connected to the larynx by the cricotracheal ligament, begins at the level of VI, the upper edge of the VII cervical vertebra and ends at the level of the upper edge of the V thoracic vertebra, where it divides into two main bronchi.
The length of the trachea varies from 8.5 to 15 cm, more often it is 10-11 cm. The trachea is a tube, somewhat compressed in the anteroposterior direction, therefore its transverse size (15-18 mm) is slightly larger than the sagittal size (14-16 mm) . The thyroid gland is adjacent to the cervical part of the trachea, the isthmus of which covers the trachea in front at the level of the second to fourth tracheal cartilages. In front, the trachea is covered by the pretracheal plate of the cervical fascia and the paired sternohyoid and sternothyroid muscles lying in it. Behind the trachea is the esophagus, and on the sides of it are the neurovascular bundles of the neck (common carotid artery, internal jugular vein and vagus nerve). Adjacent to the thoracic part of the trachea in front are the aortic arch, the brachiocephalic trunk, the left brachiocephalic vein, the beginning of the left common carotid artery and the thymus. The mucous membrane
of the trachea is lined with pseudostratified multirow columnar (cylindrical) ciliated epithelium lying on the basement membrane.
The lamina propria of the mucous membrane
contains longitudinally oriented elastic fibers, lymphocytes, lymphoid nodules and individual circularly arranged bundles of smooth myocytes.
The submucosa
gradually transforms into the dense fibrous connective tissue of the perichondrium of the tracheal cartilage.
The submucosa contains mixed serous-mucosal glands, which predominate in the posterior and lateral walls of the trachea. The excretory ducts of the glands expand in a flask-like manner and open on the surface of the mucous membrane. The fibromuscular-cartilaginous membrane of the trachea
is formed by 16-20
hyaline cartilages (cartilagines tracheales),
each of which is an arch, open posteriorly, occupying approximately two-thirds of the circumference of the trachea.
The trachea, due to the presence of cartilage in its wall, connected by dense fibrous tissue, and the membranous part, is very elastic and elastic. The trachea resists significant external pressure, its lumen remains constantly open. The trachea is covered with an adventitial membrane , consisting of loose fibrous unformed connective tissue. Blood supply to the trachea.
The trachea is supplied with blood by tracheal branches (from the inferior thyroid, internal thoracic arteries and from the aorta).
The arteries branch out and form a capillary network under the epithelium. Venous blood flows through the veins of the same name into the right and left brachiocephalic veins. Lymphatic vessels
are directed to the deep cervical lateral (internal jugular), pre- and paratracheal, upper and lower tracheobronchial lymph nodes.
innervated
by the tracheal branches of the recurrent laryngeal nerve (sensory innervation) and from the sympathetic trunk (efferent innervation).
Age-related features of the trachea. The beginning of the trachea in infants lies high, at the level of the IV-V cervical vertebrae, in adults - at the level of the VI cervical vertebra, in the elderly it descends to the VII cervical vertebra.
In women, the beginning of the trachea lies slightly higher than in men. When swallowing or moving the head, the position of the upper end of the trachea changes. In children under the age of 1 year, the tracheal bifurcation is located at the level of the III thoracic vertebra, from 2 to 6 years - at the level of IV-V, from 7 to 12 years - at the level of the V-VI thoracic vertebrae. In a newborn, the length of the trachea is 3.2-4.5 cm. The width of the lumen in the middle part does not exceed 0.8 cm. The membranous wall of the trachea is relatively wide, the tracheal cartilage is thin and soft. After birth, the trachea grows rapidly during the first 6 months, then its growth slows down and accelerates again during puberty (12-25 years). By the age of 3-4 years of a child’s life, the width of its lumen increases by 2 times. The bronchi branch out to form the so-called bronchial tree. The bronchial tree consists of bronchi of many orders of branching, the lumen of which gradually decreases. The beginning of the bronchial tree in each lung is the main bronchi, formed as a result of the bifurcation of the trachea. Right main bronchus (bronchus principalis dexter)
wider and shorter than the left one, it is almost a continuation of the trachea.
superior lobar bronchus (bronchus lobaris superior dexter)
departs from and goes to the upper lobe of the right lung; the right pulmonary artery passes under it.
The middle lobar
and
lower lobar bronchus (bronchus lobaris medius dexter
et
bronchus lobaris inferior dexter)
are directed to the corresponding lobes of the right lung.
The skeleton of the right main bronchus consists of 6-8 cartilaginous half-rings, the left main bronchus - of 9-12 half-rings. The left main bronchus (bronchus principalis sinister)
is immediately divided into
the upper
and
lower lobar (bronchus lobaris superior sinister
et
bronchus lobaris inferior sinister),
which are included in the same lobes of the left lung.
The aortic arch bends through the left main bronchus, and the azygos vein bends through the right. Secondary, or lobar, bronchi depart from the main bronchi, giving rise to smaller tertiary (segmental) bronchi, which subsequently branch dichotomously. The cross-sectional area of a branching bronchus is less than the sum of the cross-sectional areas of its branches. The international classification provides the name and numbering of each bronchopulmonary segment. The division of the bronchi, their name and numbering are presented in Table. 9. Segmental bronchi are divided into subsegmental
(1, 2, 3 generations, 9-10 in total),
lobular, intralobular
.
The bronchi are lined with pseudostratified (multi-row) columnar (cylindrical) ciliated epithelium, the thickness of which decreases as the caliber of the bronchus decreases. The thin subepithelial basement membrane
consists of a basal lamina overlying a thin layer of reticular fibrils.
The lamina propria of the mucous membrane
consists of two layers.
The thin inner layer of connective tissue is rich in collagen fibers, lymphocytes and other migrated blood cells, blood capillaries. The outer connective tissue layer of the lamina propria contains a large number of longitudinally oriented elastic fibers. Large bronchi have a continuous elastic membrane.
As their caliber decreases in the lamina propria of the mucous membrane, the number of longitudinal elastic fibers increases.
The thin muscular plate of the mucous membrane
consists of intersecting circular bundles of smooth myocytes.
In small bronchi (up to 1-2 mm in diameter), the muscular plate of the mucous membrane becomes relatively thicker. The submucosa
is formed by loose fibrous connective tissue, in which numerous mixed glands are located, pouring their secretions into the lumen of the bronchi.
The hyaline cartilages
of the main bronchi are arches, open posteriorly, where their ends are connected by the membranous part.
The cartilages are connected to each other by annular ligaments, similar to tracheal ligaments. As the caliber of the bronchi decreases, the cartilage gradually changes shape, forming cartilaginous plates of unequal shape and size, connected to each other by a dense fibrous membrane. In small bronchi, cartilaginous plates lie in the outer (adventitial
) membrane.
Cartilage completely disappears in bronchioles with a diameter of about 1 mm. In the walls of the main bronchi, muscle tissue is located in the same way as in the trachea. In the walls of the intrapulmonary bronchi there is a circular layer of smooth muscle cells located between the mucous membrane and cartilage. The diameter of the smallest branches of the airways - bronchioles - is from 0.5 to 1 mm. There are about 20 generations of bronchi, the last being the terminal bronchioles,
which are divided into 14-16
respiratory bronchioles
each.
Unlike the bronchi, the walls of the bronchioles lack cartilage. The mucous membrane of the bronchioles
is lined
with a single layer of single-row ciliated epithelium,
between the cells of which there are individual secretory
bronchiolar exocrinocytes (Clara cells),
which, according to modern data, are the source of restoration of the epithelium of the terminal bronchioles.
The lamina propria of the mucous membrane
is formed by loose fibrous connective tissue containing elastic and collagen fibers, fibroblasts, and lymphoid cells.
There are no glands. The muscular plate of the mucous membrane
is formed by spiral bundles of smooth myocytes, forming a continuous layer in larger bronchioles and a discontinuous layer in the terminal ones.
The thin submucosa,
in which the capillary network lies, passes into a very thin
outer (adventitial) membrane .
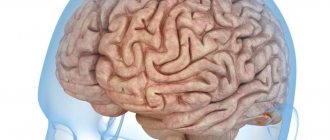
Cerebral hemispheres
1. Remember from your zoology course what parts of the brain all vertebrates have. The cerebral hemispheres are derivatives of which division? In which group of animals do they first appear?
All vertebrates are characterized by the presence of 5 parts of the brain: medulla oblongata, hindbrain, midbrain, diencephalon, forebrain.
The cerebral hemispheres are derivatives of the forebrain and first appear in amphibians, but in them they are poorly developed, and the cerebral cortex is practically absent.
2. Describe the structure of the human cerebral hemispheres. Draw a schematic cross-section of the cerebral hemispheres, indicating the gray matter of the cortex, gray matter of the nuclei, white matter, and ventricles.
In humans, the forebrain is represented by two hemispheres and the corpus callosum, which connects the hemispheres. The cerebral hemispheres: right and left - cover the midbrain and diencephalon and make up up to 80% of the mass of the adult brain. On the surface of each hemisphere there are many grooves and convolutions. There are 4 main sulci (central, lateral and parieto-occipital), which divide each hemisphere into lobes. The superficial layer consists of gray matter (cortex), underneath is white matter, consisting of axons of nerve cells whose bodies lie in the cortex or which carry information to cortical cells. In the thickness of the white matter there are large accumulations of gray matter (subcortical nuclei) and cavities (lateral ventricles). CSF circulates through the ventricles of the brain and the central canal of the spinal cord, which provides nutrition to the subcortical structures.
3. What is the cerebral cortex? Where is it?
The superficial layer of gray matter of the cerebral hemispheres is called the cortex. The cortex consists of several layers of neuron bodies, different in structure and function. It is believed that it consists of about 12-18 billion cells, the thickness is 1.5 - 4.5 mm, and the area is 1.7 - 2.5 thousand cm2.
4. Explain the significance of the grooves and convolutions on the surface of the cerebral hemispheres.
The furrows and convolutions significantly increase the surface of the cerebral cortex, according to some sources up to 10-12 times.
5. What function does the white matter of the cerebral hemispheres perform?
White matter forms pathways that connect areas of the cortex and the cortex with the rest of the nervous system.
6. What lobes are distinguished in the cerebral hemispheres?
The hemispheres are divided into frontal, 2 parietal, 2 temporal and occipital lobes
7. Distinguish between the concepts of “lobes of the cerebral hemispheres” and “zones of the cerebral hemispheres”. Give examples of when they coincide or do not coincide.
The lobes of the cerebral hemispheres are a division of the surface of the cortex according to an anatomical principle: in each hemisphere there are frontal, occipital, parietal, and temporal lobes.
Cortical zones are a section of the cerebral cortex, characterized by uniformity of structure and functions.
In our brain, the lobes and zones do not coincide; one lobe consists of several zones. The occipital lobe consists of the areas of vision and visual recognition. The areas of smell, hearing and taste are located in and adjacent to the temporal lobe.
8. When examining a blind patient, it was discovered that his eyes and optic nerves were not damaged. Why doesn't he still see?
The inability to see can be caused not only by damage to the eyes or optic nerves, but also by damage to the visual areas of the occipital lobes of the cerebral hemispheres.
9. Using additional sources of information, find out whether the functions of the left and right hemispheres of the cerebrum differ.
There is a “functional asymmetry” between the right and left hemispheres of the brain, that is, their functions are different. This was proven during experiments on cutting the communication routes between the hemispheres (subsequently such operations began to be carried out for medical reasons for certain diseases, such as Parkinson's disease). In right-handed people, the leading hemisphere is the left, in left-handed people, the leading hemisphere is the right. The right hemisphere is responsible for imaginative thinking, forms the basis for creativity and making non-standard decisions; The visual area of the right hemisphere is responsible for recognizing faces. The left hemisphere provides logical reasoning and abstract thinking, it contains the centers of oral and written speech, and the formation of decisions; The visual area is responsible for recognizing letters and numbers. Therefore, as a joke, right-handers are called mathematicians, and left-handers are called artists.
10. There are everyday concepts of “male logic” and “female logic”. Is there any basis for such differences?
In the external and internal structure, there is no difference between the brains of men and women, but despite the fact that the structure of our brains is almost the same, we are individual. The main differences in the perception of the world according to the “male” or “female” type are laid down in early childhood, depending on our upbringing and under the influence of gonadal hormones during growing up, but all of them are only psychological in nature.
Where do convolutions come from?
Scientists from the Institute of Neurobiology named after. Max Planck discovered a previously unknown mechanism leading to the formation of grooves and convolutions on the surface of the mammalian brain. The project manager was Rüdiger Klein. The results of the study were published on May 4 on the institute’s website.
The grooves and convolutions of the brain are very important because they increase the area of this organ, improving its ability to think and other actions and processes. However, not all mammals have convolutions: the brains of mice, for example, are smooth. Young neurons that migrate to the cerebral cortex during smooth brain development have so-called FLRT receptors on the surface of their cells. This provides a certain level of cohesion between cells and supports the normal movement of neurons, which leads to the formation of a smooth brain. Compared to the mouse brain, FLRTs are present in much lower abundance in the human brain. As scientists showed in this experiment, when the expression of FLRT is experimentally reduced, convolutions similar to those observed in humans begin to appear in the mouse brain. The new data represent a significant contribution to the study of brain development and formation in mammals.
Scientists have demonstrated that convolutional brains contain large numbers of progenitor cells that produce many young neurons that migrate to the cortex as the brain develops. As a result of this movement, the cellular layer of the brain is filled with neurons. By means of some mechanisms that limit the growth of the cortex, convolutions are formed so that there is enough space for the assembled cells. Experiments on mice with artificially increased numbers of progenitor cells showed that this change alone was not enough to cause gyri and sulci to appear, since in some cases the mice's cortex became thicker but remained generally smooth.
Previously, the researchers found that migrating neurons are held together by the molecule FLRT3, and they also suggested that this molecule may be involved in the process of furrow formation. To test this theory, they bred mice whose progenitor cells lacked either FLRT3 or its associated receptor, FLRT1. And although no changes were made to the number of progenitor cells, convolutions appeared on the animals' brains. Scientists explained this by saying that, due to the absence of FLRT1 and FLRT3 receptors, migrating young neurons no longer adhered as tightly as before. As a result of the action of repulsive mechanisms, clusters of neurons were formed, within which neurons moved freely and migrated faster to the surface of the brain. As a result, they reached the cortex too quickly and were distributed unevenly, which led to the formation of grooves and convolutions. In addition, this was probably facilitated by the increased softness of the bark.
In these experiments, scientists demonstrated for the first time that coupling between migrating neurons is extremely important for the formation of gyri and sulci, and FLRT receptors are essential for this process. They also demonstrated that the abundance of FLRT1 and FLRT3 receptors in humans and ferrets (which also have gyri) is significantly lower than in smooth-brained mice, suggesting that FLRT influences sulcus formation in the human brain as well. The data obtained can form the basis for subsequent studies in the field of normal and pathological development of sulci and convolutions in the mammalian brain.