Excitation processes in neuropsychiatry
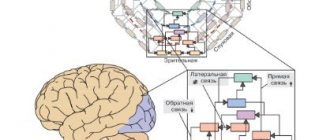
Biomarkers are necessary for diagnosis, selection and prognosis of therapy for neuropsychiatric disorders. Promising directions for the search for biomarkers are the imbalance of excitation and inhibition in the brain, as well as disorders of its maturation. Two patterns of gene expression have been identified: a decrease in markers of maturity and an increase in markers of immaturity. These two groups of genes were characterized by an overrepresentation of genes associated with synaptic function and chromosomal modification, respectively. Transcriptomic pseudoimmune disorders induced by neural hyperarousal are shared by a variety of neuropsychiatric disorders, such as schizophrenia, Alzheimer's disease and bipolar affective disorder. This endophenotype serves as the basis for transdiagnostic characterization of these neuropsychiatric disorders.
There is now evidence that patients with different clinical diagnoses share similar biological features, such as genetic mutations, molecular expression or brain activity. Recently, psychiatry has undergone a “tectonic shift” as a result of its inclusion in the concepts of modern biology. Recently, there have been repeated attempts to reclassify mental disorders according to biological domains (eg, genes, neural circuits, behavior), for example, through the Research Domain Criteria (RDoC) initiative. Therefore, identifying biomarkers that can be used for the transdiagnostic assessment of neuropsychiatric disorders is essential to better classify these diseases and understand their biological basis.
Cross-expression of genes with different disorders can be used to characterize the five major neuropsychiatric disorders. Some of these overlapping gene groups have been biologically well characterized by gene ontology enrichment or cell type specificity, but the biological properties of other gene groups remain rather unclear. Thus, unbiased coexpression network analysis does not necessarily reveal modules that highlight biological features of neuropsychiatric disorders.
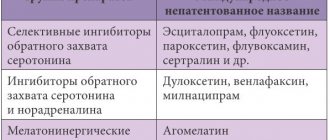
Brain pseudoimmunity can be induced in adulthood. Long-term treatment with fluoxetine was previously found to reverse the maturation status of dentate gyrus (DG) neurons in adult wild-type mice, a phenomenon that can be termed “dematuration.” Likewise, recent studies suggest that several maturity-related genes and electrophysiological properties in the DG of adult wild-type mice acquire an immature status following treatment with pilocarpine or electroconvulsive stimulation. An IDG-like phenotype has been found in patients with epilepsy. It can be assumed that neuronal hyperexcitation may be the cause of pseudo-immaturity of the brain in adulthood.
Some studies suggest that neuronal hyperexcitation may underlie abnormalities associated with certain types of neuropsychiatric disorders. Patients with epilepsy have an increased risk of developing schizophrenia, and conversely, patients with epilepsy may also exhibit psychotic symptoms that are similar to those found in patients with schizophrenia. Imbalances in excitatory and inhibitory brain circuits have been suggested to be involved in the pathogenesis of schizophrenia. Overactive action potential firing has also been observed in hippocampal granule cell-like neurons derived from induced pluripotent stem cells from patients with bipolar disorder. Recent studies have shown that patients with Alzheimer's disease and temporal lobe epilepsy may share common mechanisms underlying these neuropsychiatric disorders. Immature gene expression patterns induced by neuronal hyperexcitation may overlap with pathological gene expression patterns in the brains of patients with neuropsychiatric disorders.
Many genetic risks and environmental factors, such as seizures, hypoxia and infection, contribute to the development of neuropsychiatric disorders. Many of the data sets from patients with schizophrenia and from the postmortem brains of patients with Alzheimer's disease showed changes in the structure of the dominant hiM index. hiM genes include the GABA receptor, voltage-gated calcium channel, glutamate receptor, and voltage-gated sodium channel. These genes have been reported to be involved in pathological changes in the brains of patients with schizophrenia and Alzheimer's disease. Thus, many of the synaptic genes that are altered in the brains of patients with schizophrenia or Alzheimer's disease may be genes whose expression increases during maturation and decreases when the nervous system is hyperexcited. Although decreased expression of some synaptic genes in these disorders is well documented, a number of studies point to the possibility that neuronal hyperexcitation may also cause contractions in such synaptic molecules.
Most data sets from patients with bipolar affective disorder and Alzheimer's disease showed dominant hiI patterns. hiI genes include DNA methyltransferase, cyclin, cyclin-dependent kinase, integrin beta-3 binding protein, and tumor protein p53. These genes are known to play important roles in chromosomal modification and DNA repair, and abnormal functions of these systems have been observed in patients with bipolar disorder and Alzheimer's disease. Thus, some of the genes thought to be important in the development of these disorders are immaturity-associated genes whose expression decreases during maturation and can be increased by neuronal hyperexcitation.
Regarding datasets from patients with Parkinson's disease (PD), bipolar disorder (BPD), Huntington's disease (HD), and major depressive disorder (MDD), most of them did not have significant overlap in either the hiM or hiI genes, indicating that there cannot be pathological changes in pseudoimmunity induced by nervous system hyperexcitation in the data sets of these four diseases. Thus, it can be assumed that gene expression analysis based on the results of common endophenotypes is useful for transdiagnostic characterization of neuropsychiatric disorders.
Excitation
(biological), the reaction of a living cell to irritation, developed in the process of evolution.
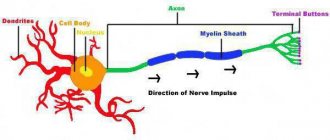
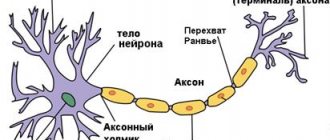
With V., the living system moves from a state of relative physiological rest to activity (for example, contraction of muscle fibers, secretion of secretions by glandular cells, etc.). V. is based on complex physical and chemical processes. The initial starting moment of V. is a change in the ionic permeability and electrical potentials of the membrane. V. has been most fully studied in nerve and muscle cells, where it is accompanied by the appearance of an action potential (AP), which can spread along the entire cell membrane without attenuation (non-decremental) (see Biological membranes
).
This property of the PD ensures the rapid transmission of information along the peripheral nerves to the nerve centers and from them to the executive organs - muscles and glands. In skeletal muscle fibers, the action force propagates both along the membrane and deep into the fiber to the contractile apparatus of the myofibrils; therefore, following the V wave, a contraction wave propagates along the muscle fiber. PD also has a triggering effect on the secretion by nerve endings of chemical substances - mediators
that have an exciting or inhibitory effect on innervated tissues.
AP obeys the “all or nothing”
: it occurs only after the stimulus reaches a threshold value (irritation threshold) and immediately acquires maximum amplitude.
During the development of AP, the cell completely loses excitability
, i.e., the ability to respond with new excitation to a repeated stimulus.
Excitability is restored gradually only after the end of the AP (see Refractory period
).
A spreading impulse is not the only form of energy. In certain areas of the nerve and muscle cells of vertebrates and in some cells of invertebrates, energy has a local character. Among the varieties of local V., the most important functional significance is the generator potentials of receptors and excitatory postsynaptic potentials that arise in the area of contact of the cell with motor nerve endings (see Synapses
).
Just like PD, local V. is associated with a selective increase in the ionic permeability of the membrane and manifests itself in the form of a negative fluctuation in surface potential (see Depolarization
). However, unlike PD, local V. does not obey the “all or nothing” rule: it has no threshold and varies in amplitude and duration depending on the strength and duration of the stimulus. The depolarization of the membrane that accompanies local V. serves as an irritant for neighboring areas of the membrane capable of generating a spreading AP. Therefore, when the local voltage (generator or postsynaptic potential) reaches a threshold value, an AP occurs. Differences in the properties of local and spreading V. are important for the processes of information transmission by nerve cells and their fibers. Local V. is characteristic of those parts of the cell membrane that are specialized to perceive stimuli coming from the outside (receptor membrane) or from other nerve cells (postsynaptic membrane). Local V. has a gradual character and therefore can more subtly reflect the characteristics of the stimulus—its strength, duration, rate of increase and decrease—than AP, the occurrence of which serves only as a signal that the stimulus has reached a threshold value. On the other hand, the ability of PD to rapidly spread without decrement makes it most adequate for transmitting information over long wires. In this case, information about the strength, duration, and steepness of changes in the stimulus is encoded by the frequency of nerve impulses, the change in this frequency over time, and the duration of the entire AP burst.
The mechanism of generation and conduction of nerve impulses, improving in the process of evolution of organisms, reached its highest development in warm-blooded animals and humans. The speed of conduction of blood in the pulpy (myelinated) fibers of warm-blooded animals reached 100-120 m/sec
with an AP duration of 0.2–0.4
m/sec
.
In cold-blooded vertebrates (for example, amphibians), the speed of AP conduction through pulp fibers of the same diameter (20 μm
) does not exceed 20-30
m/sec
with an AP duration of 1.5-2
m/sec
.
V. is a complex reaction in which, along with electrical components, structural, chemical (including enzymatic), physical (temperature), and other components also play a significant role. Changes in the membrane potential during V. are a consequence of a selective increase in the permeability of the membrane (as a result of its molecular rearrangement) for extracellular and intracellular ions (see Membrane theory of excitation
).
It is assumed that this is also associated with increased heat production during the ascending phase of PD and heat absorption during the descending phase. The penetration of Na+ and (or) Ca2+ ions into the protoplasm during V. activates enzymatic processes aimed at restoring the initial inequality of concentrations of Na+, K+, Ca2+ ions on both sides of the membrane (see “ Sodium pump”
) and at the synthesis of proteins and phospholipids, necessary for the constant renewal of the structure of the membrane and protoplasm of the cell.
Activation of metabolic processes is accompanied by an increase in oxygen consumption by the tissue and a new increase in heat production, which lasts in the nerve for many minutes after the passage of the impulse. Wed. Braking
.
Lit.:
Human Physiology, ed. E. B. Babsky, 2nd ed., M., 1936; Eccles J., Physiology of Nerve Cells, trans. from English, M., 1959; Hodgkin A., Nerve impulse, trans. from English, M., 1965; Katz B., Nerve, muscle, synapse, trans. from English, M., 1968; Khodorov B.I., The problem of excitability, Leningrad, 1969.
B. I. Khodorov.
Table of contents