Нервные клетки или нейроны представляют собой электрически возбудимые клетки, которые обрабатывают и передают информацию с помощью электрических импульсов. Такие сигналы передаются между нейронами через синапсы. Нейроны могут связываться друг с другом в нейронные сети. Нейроны являются основным материалом головного и спинного мозга центральной нервной системы человека, а также ганглий периферической нервной системы человека.
Нейроны бывают нескольких типов в зависимости от функций:
- Сенсорные нейроны, реагирующие на такие раздражители как свет, звук, прикосновения, а также на другие стимулы, влияющие на клетки органов чувств.
- Двигательные нейроны, посылающие сигналы к мышцам.
- Интернейроны, соединяющие одни нейроны с другими в головном мозге, спинном мозге или в нейронных сетях.
Строение нейрона
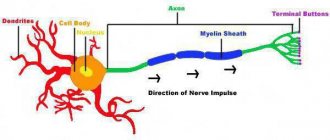
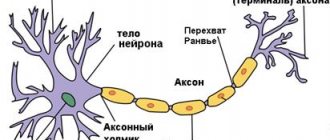
Обычный нейрон состоит из тела клетки (сомы), дендритов и аксона. Дендриты представляют собой тонкие структуры, идущие от тела клетки, они имеют многоразовое ветвление и размеры в несколько сотен микрометров. Аксон, который в миелизированном виде ещё называют нервным волокном, представляет собой специализированное клеточное расширение, берущее начало из тела клетки из места под названием аксоновый холмик (бугорок), распространяется на расстояние до одного метра. Часто нервные волокна связываются в пучки и в периферическую нервную систему, формируя нервные нити.
Содержащая ядро цитоплазматическая часть клетки называется телом клетки или сомой. Обычно, тело каждой клетки имеет размеры от 4 до 100 мкм в диаметре может быть различных форм: веретенообразной, грушевидной, пирамидальной, а также намного реже звездообразной формы. Тело нервной клетки содержит большое сферическое центральное ядро с множеством гранул Ниссля с цитоплазматической матрицей (нейроплазмой). Гранулы Ниссля содержат в себе рибонуклеопротеид и принимают участие в синтезе белка. Нейроплазма также содержит в себе митохондрии и тела Гольджи, меланин и липохромные пигментные гранулы. Количество данных клеточных органелл зависит от функциональных особенностей клетки. Следует отметить, что тело клетки существует с нефункциональной центросомой, что не даёт нейронам возможности делиться. Вот почему количество нейронов у взрослого человека равно количеству нейронов при рождении. По всей длине аксона и дендритов присутствуют хрупкие цитоплазматические нити, называемые нейрофибриллами, берущие своё начало от тела клетки. Тело клетки и её придаток окружены тонкой мембраной под названием нейронная мембрана. Описанные выше клеточные тела присутствуют в сером веществе головного и спинного мозга.
Короткие цитоплазматические придатки тела клетки, получающие импульсы от других нейронов называются дендритами. Дендриты проводят нервные импульсы в тело клетки. Дендриты имеют начальную толщину от 5 до 10 мкм, но постепенного их толщина уменьшается и они продолжаются обильным ветвлением. Дендриты получают импульс от аксона соседнего нейрона через синапс и проводят импульс к клеточному телу, поэтому их называют рецептивными органами.
Длинный цитоплазматический придаток клеточного тела, передающий импульс от тела клетки к соседнему нейрону называется аксон. Аксон значительно превышает размеры дендритов. Аксон берет своё начало в конической высоте тела клетки, называемым аксоновым холмиком, лишенным гранул Ниссля. Длина аксона является переменной и зависит от функционально связи нейрона. Цитоплазма аксона или аксоплазма содержит нейрофибриллы, митохондрии, но в ней нет гранул Ниссля. Мембрана, которая покрывает аксон имеет название аксолемма. Аксон может давать отростки, называемые добавочными вдоль своего направления, а ближе к концу аксон имеет интенсивное ветвление, заканчивающееся кистью, последняя его часть имеет увеличение для формирования бульбы. Аксоны присутствуют в белом веществе центральной и периферической нервной системы. Нервные волокна (аксоны) покрыты тонкой мембраной, которая богата липидами и называется миелиновой оболочкой. Миелиновая оболочка сформирована шванновскими клетками, которые покрывают нервные волокна. Часть аксона, не покрытая миелиновой оболочкой представляет собой узел смежных миелинизированных сегментов называемым узлом Ранвье. Функция аксона заключается в передаче импульса из клеточного тела одного нейрона в дендрон другого нейрона через синапс. Нейроны специально предназначены для передачи межклеточных сигналов. Разнообразие нейронов связано с выполняемыми ими функциями, размеры сомы нейронов варьируются от 4 до 100 мкм в диаметре. Ядро сомы имеет размеры от 3 до 18 мкм. Дендриты нейрона представляют собой клеточные придатки, образующие целые дендритные ветви.
Аксон является самой тонкой структурой нейрона, но его длина может превышать диаметр сомы в несколько сот и тысяч раз. Аксон несёт нервные сигналы от сомы. То место, где из сомы выходит аксон называется аксоновым холмиком. Длина аксонов может быть разной и достигает в некоторых участках организма длины более 1 метра (например от основания позвоночника до кончика пальца ноги).
Между аксонами и дендритами существуют некоторые структурные различия. Так, типичные аксоны практически никогда не содержат рибосомы, за исключением некоторых в начальном сегменте. Дендриты содержат гранулированный эндоплазматический ретикулум или рибосомы, уменьшающийся с удалением от тела клетки.
Мозг человека имеет очень огромное количество синапсов. Так, каждый из 100 миллиардов нейронов содержит в среднем 7 000 синаптических связей с другими нейронами. Установлено, что мозг трехлетнего ребёнка имеет около 1 квадриллиона синапсов. Количество этих синапсов уменьшается с возрастом и стабилизируется у взрослых. У взрослого количество синапсов составляет от 100 до 500 триллионов. Согласно исследованиям мозг человека содержит около 100 миллиардов нейронов и 100 триллионов синапсов.
Структурная классификация нейронов
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
· Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
· Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
· Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
· Мультиполярные нейроны — Нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе
· Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (т. е. находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация нейронов По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние – неультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — эта группа нейронов осуществляет связь между эфферентными и афферентными, их делят на комиссуральные и проекционные (головной мозг).
Морфологическая классификация нейронов Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
1. учитывают размеры и форму тела нейрона,
2. количество и характер ветвления отростков,
3. длину нейрона и наличие специализированные оболочки.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120-150 мкм у гигантских пирамидных нейронов. Длина нейрона у человека составляет от 150 мкм до 120 см. По количеству отростков выделяют следующие морфологические типы нейронов: — униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге; — псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях; — биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях; — мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Развитие и рост нейрона Нейрон развивается из небольшой клетки — предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным.) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему. Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона. Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне.
Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки. Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Нейроглия. В отличие от нервных клеток, глиальные клетки обладают большим разнообразием. Их количество в десятки раз превышает количество нервных клеток. В отличие от нервных клеток, глиальные способны делиться, их диаметр значительно меньше диаметра нервной клетки и составляет 1,5-4 микрона.
Долгое время считали, что функция глиоцитов несущественна, и они выполняют лишь опорную функцию в нервной системе. Благодаря современным методам исследования, установлено, что глиоциты выполняют ряд важных для нервной системы функций: опорная, разграничительная, трофическая, секреторная, защитная.
Среди глиоцитов, по морфологической организации, выделяют ряд типов: эпендимоциты, астроциты.
Эпендимоциты образуют плотный слой клеток, элементов, выстилающих спинномозговой канал и желудочки мозга. В процессе онтогенезе, эпендимоциты образовывались из спонгиобластов. Эпендимоциты представляют собой слегка вытянутые клетки с ветвящимися отростками. Некоторые эпендимоциты выполняют секреторную функцию, выделяя биологически активные вещества в кровь и в желудочки мозга. Эпендимоциты образуют скопления на капиллярной цепи желудочков мозга; при введении в кровь красителя, он накапливается эпендимоцитах, это свидетельствует о том, что последние выполняют функцию гематоэнцефалического барьера.
Астроциты выполняют опорную функцию. Это огромное количество глиальных клеток, имеющих множество коротких отростков. Среди астроцитов выделяют 2 группы:
o плазматические клетки
o волокнистые астроциты
Олигодендроциты – крупные глиальные клетки, часто сконцентрированы вокруг нервной клетки и поэтому называются сатиллитными глиацитами. Их функция очень важна для трофики нервной клетки. При функциональных перенапряжениях нервной клетки, глиоциты способны прореферировать вещества поступающие путем пиноцитоза в нервную клетку. При функциональных нагрузках, вначале происходит истощение синтетического аппарата глиальных клеток, а затем нервных. При восстановлении (репарации), вначале восстанавливаются функции нейронов, а затем – глиальных клеток. Таким образом, глиоциты принимают участие в обеспечении функций нейронов. Глиальные клетки существенным образом способны влиять на трофику мозга, а также на функциональный статус нервной клетки. Нервы (nervi) — анатомические образования в виде тяжей, построенные преимущественно из нервных волокон и обеспечивающие связь центральной нервной системы с иннервируемыми органами, сосудами и кожным покровом тела.
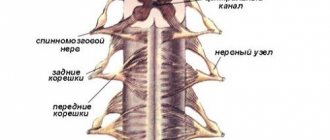
Нервы отходят парами (левый и правый) от головного и спинного мозга. Различают 12 пар черепных нервов и 31 пару спинномозговых Н.; совокупность Н. и их производных составляет периферическую нервную систему (рис.), в составе которой в зависимости от особенностей строения, функционирования и происхождения выделяют две части: соматическую нервную систему, иннервирующую скелетные мышцы и кожный покров тела, и вегетативную нервную систему,
иннервирующую внутренние органы, железы, кровеносную систему и др.
Нервные волокна — отростки нервных клеток (нейронов), имеющие оболочку и способные проводить нервный импульс. Главной составной частью нервного волокна является отросток нейрона, образующий как бы ось волокна. Большей частью это аксон. Нервный отросток окружен оболочкой сложного строения, вместе с которой он и образует волокно. Толщина нервного волокна в организме человека, как правило, не превышает 30 микрометров. Нервные волокна делятся на мякотные (миелиновые) и безмякотные (безмиелиновые). Первые имеют миелиновую оболочку, покрывающую аксон, вторые лишены миелиновой оболочки.
Как в периферической так и в центральной нервной системе преобладают миелиновые волокна. Нервные волокна, лишенные миелина располагаются преимущественно в симпатическом отделе вегетативной нервной системы. В месте отхождения нервного волокна от клетки и в области перехода его в конечные разветвления нервные волокна могут быть лишены всяких оболочек, и тогда они называются голыми осевыми цилиндрами.
В зависимости от характера проводимого по ним сигнала, нервные волокна подразделяют на двигательные вегетативные, чувствительные и двигательные соматические.
Строение нервных волокон:
Миелиновое нервное волокно имеет в своём составе следующие элементы (структуры): 1) осевой цилиндр, располагающийся в самом центре нервного волокна, 2) миелиновую оболочку, покрывающую осевой цилиндр, 3) шванновскую оболочку.
Осевой цилиндр состоит из нейрофибрилл. Мякотная оболочка содержит большое количество веществ липоидного характера, известных под названием миелина. Миелин обеспечивает быстроту проведение нервных импульсов. Миелиновая оболочка покрывает осевой цилиндр не на всём промежутке, образуя промежутки, получившие название перехваты Ранвье. В области перехватов Ранвье осевой цилиндр нервного волокна примыкает к верхней — шванновской оболочке.
Промежуток волокна, расположенный между двумя перехватами Ранвье, называют сегментом волокна. В каждом таком сегменте на окрашенных препаратах можно видеть ядро шванновской оболочки. Оно лежит приблизительно посредине сегмента и окружено протоплазмой шванновской клетки, в петлях которой и содержится миелин. Между перехватами Ранвье миелиновая оболочка также не является сплошной. В толще ее обнаруживаются так называемые насечки Шмидт-Лантермана, идущие в косом направлении.
Клетки шванновской оболочки, так же как и нейроны с отростками, развиваются из эктодермы. Они покрывают осевой цилиндр нервного волокна периферической нервной системы аналогично тому, как клетки глии покрывают нервное волокно в центральной нервной системе. В результате этого они могут называться периферическими глиальными клетками.
В центральной нервной системе нервные волокна не имеют шванновских оболочек. Роль шванновских клеток здесь выполняют элементы олигодендроглии. Безмиелиновое (безмякотное) нервное волокно лишено миелиновой обкладки и состоит только из осевого цилиндра и шванновской оболочки.
Функция нервных волокон.
Главная функция нервных волокон – передача нервного импульса. В настоящее время изучено два типа нервной передачи: импульсная и безимпульсная. Импульсная передача обеспечивается электролитными и нейротрансмиттерными механизмами. Скорость передачи нервного импульса в миелиновых волокнах значительно выше, чем в безмякотных. В её осуществлении важнейшая роль принадлежит миелину. Данное вещество способно изолировать нервный импульс, в результате чего передача сигнала по нервному волокну происходит скачкообразно, от одного перехвата Ранвье к другому. Безимпульсная передача осуществляется током аксоплазмы по специальным микротрубочкам аксона, содержащим трофогены – вещества, оказывающие на иннервируемый орган трофическое влияние.
Ганглий (др.-греч. γανγλιον — узел) или нервный узел — скопление нервных клеток, состоящее из тел, дендритов и аксоновнервных клеток и глиальных клеток. Обычно ганглий имеет также оболочку из соединительной ткани. Имеются у многих беспозвоночных и всех позвоночных животных. Часто соединяются между собой, образуя различные структуры (нервные сплетения, нервные цепочки и т. п.).
БИЛЕТ № 13
1. Кости лицевого черепа. Глазница. Носовая полость. Сообщения.
2. Толстая кишка: отделы, их топография, строение, отношение к брюшине, кровоснабжение и иннервация.
3. Продолговатый мозг. Внешнее и внутреннее строение. Топография серого и белого вещества.
Виды нейронов
Нейроны бывают нескольких форм и размеров и классифицируются по их морфологии функциям. Так например, анатом Камилло Гольджи разделял нейроны на две группы. К первой группе он отнёс нейроны с длинными аксонами, которые передают сигналы на длинные расстояния. Ко второй группе он относил нейроны с короткими аксонами, которые можно было спутать с дендритами.
Нейроны классифицируются по своему строению на следующие группы:
- Однополярные. Аксон и дендриты выходят из одного придатка.
- Биполярные. Аксон и одиночный дендрит располагаются на разных сторонах сомы.
- Многополярные. Не менее двух дендритов располагаются отдельно от аксона.
- Тип Гольджи I. Нейрон имеет длинный аксон.
- Тип Гольджи II. Нейроны, у которых аксоны расположены локально.
- Анаксонные нейроны. Когда аксон не отличим от дендритов.
- Корзинные клетки — интернейроны, формирующие плотно сплетённые окончания по всей соме клеток-мишеней. Присутствуют в коре головного мозга и в мозжечке.
- Клетки Беца. Представляют собой большие двигательные нейроны.
- Клетки Люгаро — интернейроны мозжечка.
- Средние остроконечные нейроны. Присутствуют в полосатом теле.
- Клетки Пуркинье. Представляют собой крупные многополярные нейроны мозжечка типа Гольджи I.
- Пирамидальные клетки. Нейроны с сомой треугольной формы типа Гольджи II.
- Клетки Реншоу. Нейроны, с обеих концов связанные с альфа моторными нейронами.
- Униполярный кистевидные клетки. Интернейроны, которые имеют уникальные дендритные окончания в виде кисти.
- Клетки переднего роговидного отростка. Представляют собой мотонейроны, расположенные в спинном мозге.
- Шпиндельные клетки. Интернейроны, соединяющие отдаленные области мозга.
- Афферентные нейроны. Нейроны, которые передают сигналы от тканей и органов в центральную нервную систему.
- Эфферентный нейроны. Нейроны, передающие сигналы от центральной нервной системы к эффекторным клеткам.
- Интернейроны, подключающие нейроны в конкретных областях центральной нервной системы.
ВВЕДЕНИЕ В КОГНИТИВНЫЕ НЕЙРОНАУКИ. Глава 3. Нейроны и связи между ними.
ВВЕДЕНИЕ В КОГНИТИВНЫЕ НЕЙРОНАУКИ. Из учебника МОЗГ. ПОЗНАНИЕ. РАЗУМ. Электронная книга https://t.me/kudaidem/1879
Глава 3. Нейроны и связи между ними.
Что мы знаем о процессах, происходящих на уровне нейронов? Можем ли мы сейчас построить непротиворечивую теорию относительно событий на этом уровне?
Основными клетками мозга являются нейроны, высококонсервативные с эволюционной точки зрения. Они сохранялись в относительно неизменном виде в течение многих сотен миллионов лет, и даже очень разные виды животных имеют одинаковые типы нейронов. Со многих точек зрения нейроны не отличаются от остальных клеток, но есть то, что выделяет их среди остальных: специализация на электрохимической сигнализации, благодаря которой они способны принимать входящий сигнал на дендритах и посылать электрохимический сигнал вдоль аксона. Весь мозг можно рассматривать как сверхсложную структуру, состоящую из связанных между собой нейронов.
(рис. 3.1).
Дендритами и аксонами называют выросты тела нейрона; один нейрон может иметь до десяти тысяч дендритов и один аксон.
(рис. 3.2 и 3.3).
Потенциал действия (ПД) проходит по аксону значительно медленнее, чем электрический ток в компьютере, однако многие задачи наш мозг выполняет гораздо лучше современных компьютеров. В настоящее время компьютеры далеко отстоят от человека в задачах восприятия, языковой коммуникации, семантической памяти, контроля движения и творчества.
Нейробиология концентрирует внимание на связи и взаимодействии нейронов. Рассмотрение таких связей удобно начинать как раз с генерализованного нейрона.
Классические нейроны соединяются при помощи синапсов, которые могут быть возбуждающими и тормозящими.
(рис. 3.6)
Активность нейрона опосредована десятками факторов — циклом сна и бодрствования, доступностью предшественников нейромедиаторов и многими другими. Все эти факторы влияют на вероятность прохождения сигнала между двумя нейронами и могут быть представлены в виде синаптических весов. Таким образом, все разнообразие нейронов можно с успехом представить в виде интегративного нейрона, а все способы межнейронной коммуникации — в форме вероятности прохождения сигнала между нейронами.
Существует по меньшей мере шесть основных нейромедиаторов и не менее тридцати «менее важных», в основном являющихся нейропептидами.
Даже дендриты отдельной клетки, по всей видимости, способны к обработке информации. Имеются также данные, что способна принимать участие в обработке информации и нейроглия — поддерживающая ткань нервной системы.
На настоящий момент известно о существовании в некоторых частях взрослого головного мозга стволовых клеток. Образование новых синапсов идет в течение всей жизни; для образования новых синапсов отростки дендритов способны образовываться за несколько минут.
(рис. 3.7).
1.3. Обработка информации нейронами.
Искусственные нейронные сети использовались для моделирования многих функций мозга — распознавания элементов изображений, управления роботами, обучения и улучшения функционирования на основе опыта.
Во многих случаях такие сети выполняли задачи лучше, чем компьютерные программы, основанные на логике и математике.
Так же, они помогают нам понять принципы работы реальных нейронных сетей в мозге.
Нейросети помогают нам понять работу нервной системы.
Так искусственные нейросети могут служить моделями для изучения реальных структур в мозге.
Мы ограничимся рассмотрением синапсов только двух типов — возбуждающего (повышающего вероятность прохождения ПД (Потенциал действия) на постсинаптическом нейроне) и тормозящего (понижающего такую вероятность).
Глутамат — наиболее распространенный медиатор в ЦНС — является возбуждающим.
ГАМК (гаммаАминоМасляная кислота) является наиболее распространенным тормозным медиатором.
В нервной системе распространены массивы нейронов, часто именуемые картами.
2.1. Упрощенный случай: рецепторы, пути и контуры.
Каждый сенсорный нерв может содержать несколько параллельных каналов, каждый из которых проводит несколько различающуюся информацию. Так, зрительный тракт имеет канал передачи цвета, называемый мелкоклеточным, и канал передачи формы и размеров объекта, называемый крупноклеточным.
Точно так же соматосенсорные пути сочетают каналы передачи прикосновения, давления, боли и некоторые другие.
Большинство сенсорных волокон оканчиваются в таламусе, где они передают сигнал нейронам, оканчивающимся в коре.
(рис. 3.10 и 3.11)
Таким образом, в большинстве сигнальных путей существуют петли обратной связи — такие, как в нейросети с двумя или более слоями.
С этой точки зрения мозг представляется системой воздействующих друг на друга массивов и сетей.
Массивы нейронов представляет собой двумерную сеть нейронов.
Когда массивы соответствуют пространственной организации той или иной структуры, их называют картами.
В мозге имеет место как временное, так и пространственное кодирование, наряду со многими другими способами кодирования и обработки информации.
Пространственные карты являются наиболее наглядной формой пространственного кодирования.
(рис. 3.15 и 3.16).
Такая соматосенсорная информация, как информация о прикосновении и боли, также обрабатывается кортикальными картами. Другие чувства, такие как слух, вкус и обоняние, гораздо меньше ассоциированы с положением рецепторов, однако в слуховой коре имеется картоподобный клиновидный регион.
Таким образом, даже информация от не ассоциированных с пространством органов чувств обрабатывается массивами и картами нейронов.
Наш мозг организует огромные количества входящей информации так, чтобы отражать положение окружающих объектов. Моторная кора, как вы можете догадаться, также выглядит как непропорциональная карта скелетных мышц тела.
Главным вопросом относительно сенсорики на сегодняшний день яляется вопрос о том, как осуществляется высокоуровневая обработка воспринятой информации. И модель нейросетей предоставляет один из возможных ответов.
Мозг постоянно корректирует работу моторных систем на основании сенсорной информации и адаптирует сенсорные системы при помощи моторной активности.
Сенсорные системы можно представить в виде иерархических систем, состоящих из иерархических систем низшего порядка, начиная с рецепторов и постепенно переходя ко все более сложным объектам.
Моторные системы могут быть представлены в виде иерархической структуры противоположного направления, заканчивающиеся на мотонейронах.
Идет непрерывный обмен информацией между двумя системами в процессе цикла от восприятия до действия, начиная с наинизшего и заканчивая высшими уровнями планирования, мышления и анализа возможного развития событий
(рис. 3.20 Иерархическая система из области архитектуры).
В схеме иерархической системы обработки информации, каждый массив нейронов назван картой; карты существуют на разных уровнях, и сигнал может идти вверх, вниз и к другой карте того же уровня.
При рассмотрении электрической активности десятков миллиардов нейронов мозг поневоле начинает казаться огромным оркестром, а не одним инструментом. За сотни миллионов лет эволюции в мозге появились нейроны с самыми разными видами временного и пространственного кодирования (блок 3.1).
В путях следования информации имеется множество точек выбора, с которых она может быть направлена по нескольким различным путям или быть передана на уровень выше или ниже.
Если вернуться к ступенчатой пирамиде, то такой разветвленный путь похож на путь человека к вершине: он может достигнуть ее прямым или окружным путем.
Зрительная картина мира подвержена постоянным изменениям. Однако мозг тем не менее ведет обработку таких изменений. Животное не может позволить себе не заметить хищника, прячущегося в траве, только потому, что сейчас закат, или потому, что на него падает тень.
Для того чтобы выжить, мы должны были иметь превосходную зрительную систему.
Вот, например, кошка, выслеживающая жертву, может осматривать дерево только одним глазом, тогда как другой глаз остается пассивным. Это приводит к явлению бинокулярной конкуренции — конкуренции между зрительными входами от разных глаз.
Многие животные получают от разных глаз совершенно разные входы — такие животные, как кролики и олени, вообще не имеют участков перекрытия полей зрения, поэтому для них явление бинокулярной конкуренции невозможно.
Мозг постоянно имеет некоторые ожидания относительно встречаемых им внешних условий. Спускаясь по лестнице в темноте, мы ожидаем, что под ногой будет ступенька.
При анализе неоднозначно трактуемых объектов ожидание обусловливает выбор наиболее приемлемого варианта трактовки. Многие слова в языке имеют больше одного значения, поэтому, даже читая это, вы вынуждены иметь дело с неоднозначностями. Мозг опирается не только на входящую информацию — он имеет множество причин выбора того или иного варианта, основанных на предсказании результата и ожидании.
Селективное внимание позволяет нам динамически изменять свои сенсорные предпочтения, а долговременная память увеличивает силу синапсов, ответственных за точное восприятие.
Многие ученые считают, что всю кору целиком, наряду с сопутствующими областями, такими как таламус, следует рассматривать как одну функциональную единицу. Ее часто называют таламокортикальной системой.
(Пластичный мозг).
Одним из основных свойств поведения животных является способность адаптироваться.
Основным свойством мозга, таким образом, является приспособляемость. Однако какие изменения в структуре самого мозга приводят к такой приспособляемости?
Для этих целей гораздо лучше подходят методы визуализации структур мозга, получившие интенсивное развитие в два прошедших десятилетия.
Хотя большая часть методов визуализации ориентирована на конкретную область, подчеркивая тем самым функциональное разделение, а не интеграцию, были предприняты попытки изучения обучения как системного процесса, включающего глобальные изменения структуры и функций мозга.
Развитие технологии магнитно-резонансной томографии позволило начать изучение изменений структурных компонентов связи — трактов белого вещества — под влиянием обучения.
Как было показано, обучение жонглированию вызывает изменения как в сером, так и в белом веществе мозга.
Данные результаты стали поистине революционными, поскольку долгие годы считалось, что структура мозга неизменна.
Подобные открытия, позволяющие рассматривать мозг как функционально и структурно лабильный орган, вне всякого сомнения являются шагом вперед в нашем понимании процесса обучения.
(4.0. Адаптация и обучение массивов нейронов).
Наиболее известное правило обучения нейросетей, выражаемое в лозунге «neurons that fire together, wire together».
Нейроны, которые срабатывают вместе, соединяются вместе.
(Обучение по Хэббу).
Дональд Хэбб в 1949 г. постулировал, что ассамблеи нейронов способны обучаться благодаря усилению связей между нейронами, активирующимися при стимуляции одновременно.
В основе обучения и памяти лежит эффективность синаптической связи.
Существует множество способов воздействия на эффективность синаптической передачи. Так, два нейрона могут образовать больше синапсов, в самих синапсах может вырабатываться больше нейромедиатора, рецепторы постсинаптического нейрона могут стать эффективнее.
В обучении задействовано два типа изменений; их можно рассматривать как усиленное возбуждение и усиленное торможение.
Долговременное усиление возбудимости одного нейрона называют долговременой потенциацией.
Долговременное понижение же — долговременной депрессией. Оба события имеют место в гиппокампе.
Визуально обучение по Хэббу можно представить в виде утолщения линий между узлами сети, как в простой совокупности клеток.
Модели с третьим, скрытым слоем позволяют нейросети изменять силу соединений.
Классическая трехслойная прямая сеть со скрытым слоем и настраиваемой силой взаимодействий может эффективно обучаться путем сопоставления выхода нейросети с желаемым выходом и подстройки силы соединений для достижения желаемого результата.
Процесс носит название обратного распространения ошибки обучения и во многом подобен отрицательной обратной связи.
Сети такого типа на сегодняшний день наиболее распространены.
В самоорганизующейся аутоассоциативной сети выход ставится в соответствие входу.
Такая стратегия полезна при распознавании паттернов, таких как звук знакомого голоса.
Самоорганизующиеся системы используются в природе для решения многих задач.
Сами организмы и их нервные системы можно рассматривать как самоорганизующиеся системы.
Самоорганизующаяся сеть способна справляться с фундаментальой проблемой распознавания человеческих лиц.
Человек учится реагировать на нормальные, недеформированные лица в очень ранний период жизни и вскоре становится способен отличать знакомые лица от незнакомых.
Задача, решаемая сетью, гораздо проще решаемой человеком, поскольку в модели происходит только формирование цепи.
Сеть способна обучаться предугадывать расположение рта в нижней части рисунка и двух глаз — в верхней.
4.2. Дарвинистский подход в нервной системе: выживают клетки и синапсы, наиболее приспособленные к данной задаче.
Нейральный дарвинизм предполагает, что нейроны развиваются и соединяются друг с другом в соответствии с дарвинистскими принципами.
Селекционизм — эффективный способ адаптации.
Отбор нейронов приводит к образованию долгоживущих нейронных совокупностей, выполняющих задачи адаптации, обучения, разпознавания паттернов и им подобные.
Нейросети отличаются высоким уровнем параллельности (что означает способность производить много разных вычислений одновременно) и распределенности (способности обрабатывать информацию в разных местах с использованием разных механизмов).
Это говорит о большей близости нейросетей к биологическим способам обработки информации.
Нейронные сети довольно просто перевести на язык математических выражений.
Нейросети способны обрабатывать символьную информацию, а символы могут быть переведены в нейросети.
Обучение нейросети проявляются по мере распознавания сетью входа и отсечения ею альтернативных вариантов.
Существует масса способов координации работы нейронов. Одним из них являются масштабные ритмы, координирующие работу больших групп нейронов так же, как дирижер координирует игру симфонического оркестра. Если большая масса нейронов активируется одновременно, то их активность, как правило, суммируется.
Современные данные говорят в пользу гораздо более быстрых гамма- и тета-корреляций на тех частотах, на которых мозг выполняет большую часть этой работы.
Ритмы энцефалограммы на сегодняшний день считаются сигнализирующими о разных, но скоординированных процессах.
К примеру, гамма-ритмы высокой плотности считаются ассоциированными с осознанным зрительным восприятием и процессом решения простой проблемы эквивалентности.
Альфа-ритмы традиционно ассоциируются с отсутствием задач, требующих фокусировки внимания, тогда как тета-ритмы, как полагают на настоящий момент, контролируют гиппокампальную область и фронтальную кору в процессе обращения к долговременной памяти. Дельта-ритмы — сигналы глубокого сна — группируют быструю нейрональную активность с целью консолидации полученных данных.
При проектировании самолета инженеры закладывают в его конструкцию некоторую функциональную избыточность на случай выхода из строя важнейших систем. Так, если откажет один двигатель, то большая часть самолетов будет способна дотянуть до взлетно-посадочной полосы на оставшихся.
Человек и животные также обладают определенной функциональной избыточностью.
Мозга это правило тоже касается. Мозг способен работать даже после получения весьма значительных повреждений.
6.0. Заключение.
Латеральное торможение является распространенной стратегией для выделения различий между двумя однородными областями сигнала, такими как темные пятна на светлом фоне.
Клетки сенсорных систем имеют так называемые рецептивные поля, настроенные на определенные параметры входа, такие как ориентация линии, цвет, движение, форма и тип объекта. При повышении уровня визуальных карт их разрешение падает, в то время как способность к интеграции информации растет.
Поскольку сенсорные и моторные системы изучают отдельно друг от друга, мозг представляется нам огромным сенсомоторным органом, делающим возможным непрерывные высокоуровневые взаимодействия между входом и выходом.
Пространственные массивы нейронов делают возможным пространственное кодирование, однако не стоит забывать о том, что в нервной системе имеется еще и кодирование временное. Основные ритмы энцефалограммы, как полагают, отвечают за временную координацию активности больших групп нейронов.
Последние исследования позволяют предположить, что гамма-ритм ответственен за интеграцию сенсорной информации осознанные ощущения, а тета-ритм — за извлечение информации из долговременной памяти.
Контрольные задания к этой главе.
1. Опишите основные функции интегративного нейрона.
2. Что такое латеральное торможение и какую роль оно играет в сенсорных системах?
3. Каким образом сенсорные и моторные системы можно рассматривать в форме иерархических структур?
4. Опишите роль двусторонних взаимодействий в функционировании мозга.
5. Что такое дарвинистский подход к нервной системе и какие аспекты процессов, происходящих в мозге, он затрагивает?
6. Назовите три наиболее общих свойства сенсорных систем.
Действие нейронов
Все нейроны являются электрически возбудимыми и поддерживают напряжение на своих мембранах с помощью метаболически проводимых ионных насосов, сочетающихся с ионными каналами, которые встроены в мембрану для генерации ионных дифференциалов, таких как натрий, хлорид, кальций и калий. Изменения напряжения в кросс-мембране приводит к изменению функций вольт-зависимых ионных калом. При изменении напряжения в достаточно большом уровне электрохимический импульс вызывает генерацию активного потенциала, который быстро перемещается вдоль клеток аксона, активируя синаптические связи с другими клетками.
Большинство нервных клеток являются базовым типом. Определенный стимул вызывает электрический разряд в клетке, разряд подобный разряду конденсатора. Это продуцирует электрический импульсы равный примерно 50-70 милливольтам, который называется активным потенциалом. Электрический импульс распространяется по волокну, по аксонам. Скорость распространения импульса зависит от волокна, это примерно в среднем десятки метров в секунду, что заметно ниже скорости распространения электричества, которая равна скорости света. Как только импульс достигает пучка аксона, он передаётся на соседние нервные клетки под действием химического медиатора.
Нейрон действует на другие нейроны выпуская нейротрансмиттер, связывающийся с химическими рецепторами. Эффект от постсинаптического нейрона определяется не пресинаптическим нейроном или нейротрансмиттером, а типом активируемого рецептора. Нейротрансмиттер представляет собой как бы ключ, а рецептор замок. При этом один ключ может быть использован для открытия «замков» разного типа. Рецепторы в свою очередь классифицируются на возбуждающие (увеличивающие скорость передачи), ингибирующие (замедляющие скорость передачи) и модулирующие (вызывающие длительные эффекты).
Связь между нейронами осуществляется через синапсы, в этом месте находится окончание аксона (аксоновый терминал). Нейроны, такие например как клетки Пуркинье в мозжечке могут иметь более тысячи дендритных переходов, осуществляя связь с десятками тысяч других нейронов. Другие нейроны (крупные нейронные клетки супраоптического ядра) имеют лишь один или два дендрита, каждый из которых получает тысячи синапсов. Синапсы могут быть как возбуждающими так и ингибирующими. Некоторые нейроны связываются между собой посредством электрических синапсов, которые являются прямыми электрическими соединениями между клетками.
В химическом синапсе при достижении потенциалом действия аксона, происходит открытие напряжения в кальциевом канале, что позволяет ионам кальция проникнуть в терминал. Кальций заставляет синаптические пузырьки, наполненные молекулами нейромедиаторами проникать в мембрану, высвобождая содержимое в синаптическую щель. Происходит процесс диффундирования медиаторов через синаптическую щель, которые в свою очередь активируют рецепторы на постсинаптическом нейроне. Кроме того, высоко цитозольный кальций в терминале аксона вызывает усвоение митохондриального кальция, который в свою очередь активирует митохондриальный энергетический метаболизм для производства АТФ, что поддерживает непрерывную нейротрансмиссию.
Как они работают вместе и чем отличаются
Афферентные нейроны обычно имеют два аксона, которые передают электрохимические сигналы в позвоночный столб или мозг. Оказавшись там, сигнал проходит через сеть интернейронов и через эфферентный нейрон. Афферентно-эфферентные пары нейронов, которые проходят через позвоночник, управляют рефлексами (такими, как реакция коленного рефлекса).
Появились вопросы по этой теме? Задай вопрос преподавателю и получи ответ через 15 минут! Задать вопрос
Афферентные нейроны предназначены для реагирования на различные раздражители. Например, афферентный нейрон, предназначенный для реакции на тепло, обнаруживает избыточное тепло и посылает импульс через центральную нервную систему. Затем эфферентный нейрон заставляет мышцы сокращаться, чтобы отвести тело от жары. Кожа имеет сенсорные рецепторы для тепла, холода, удовольствия, боли и давления.
Афферентные нейроны имеют круглые и гладкие клеточные тела, в то время как эфферентные нейроны имеют спутниковые тела. Афферентные нейроны обнаруживаются в периферической нервной системе, а эфферентные нейроны располагаются в центральной нервной системе. Аксоны в афферентных нейронах движутся от ганглиев (скопление нервных клеток, в которых находятся афферентные и эфферентные нейроны) к спинному мозгу. Длинный аксон фактически связан с эфферентным нейроном.
Афферентные нейроны имеют один длинный миелинизированный дендрит, тогда как эфферентные нейроны имеют более короткие дендриты. Дендрит в афферентном нейроне — это то, что отвечает за передачу нервных импульсов от рецепторов к телу клетки, в то время как в эфферентном нейроне импульсы проходят через дендрит и выходят через нервно-мышечное соединение, которое образуется между эффекторами и аксоном.