Морфология нервной системы — один из сложных разделов анатомии.
Нервная система обеспечивает взаимодействие всех органов и систем человека, осуществляет обмен информацией, определяет адаптационные взаимоотношения организма с окружающей средой.
Правильное усвоение анатомии нервной системы определяет материалистическое формирование понятия организма как целостной системы, создает базу для изучения теоретических и клинических дисциплин (невропатологии, нейрохирургии, психиатрии и др.).
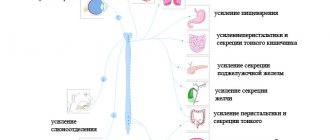
Нервная система состоит из центральной и периферической. Периферическую нервную систему образуют 12 пар черепных нервов и 31 пара спинномозговых нервов.
Изучение периферической нервной системы имеет большое практическое и диагностическое значение. В клинической практике часто встречаются такие заболевания спинномозговых нервов, как радикулит, менингорадикулит, радикулоневрит, фуникулит, нейродермит, неврит, нейромиозит, опоясывающий лишай и др.
Данная информация является дополнением к существующим анатомическим руководствам и учебникам, не заменяя их.
Функция нервной системы
— сложна и многообразна. Нервная система (system nervosum) осуществляет обмен информацией между организмом и внешней средой, регулирует и координирует функции всех органов, обеспечивает функциональное единство и целостность организма, определяет адаптивное поведение организма в окружающей среде.
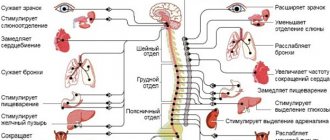
Условно нервная система делится по функциональному и топографическому принципам следующим образом (рис. 5.1).
Рис. 5.1. Схема разделения нервной системы по функциональному и топографическому принципам
Распространение нервных импульсов
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны
. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых
дендритами
, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется
аксоном
и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия
. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом
. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (
пресинаптическими
)
пузырьками
, в каждом из которых находятся особые соединения —
нейромедиаторы
. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом
). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Биохимия передачи нервного импульса. Основные компоненты и этапы.
Синапс — это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек — нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие — синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой.
АТФазы — кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза, и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц: 2-, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика — около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора. При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса «ацетилхолин-белок-рецептор» и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40 мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы.
Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые — в качестве боевых отравляющих веществ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания. Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника.
Катехоламины: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза — тирозингидроксилаза, ингибируемая конечными продуктами.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок — -ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов.
Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате изменяется генерация нервных импульсов постсинаптической мембраной (тормозится). В некторых случаях непосредственной причиной этого является повышение проницаемости постсинаптической мембраны для калия, либо снижением проводимости для натрия (эти события приводят к гиперполяризации).
ГАМК — тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала.
Глицин — тормозной медиатор, по вызываемым эффектам подобен гамк.
Образование нейромедиаторов – ацетилхолина, адреналина, дофамина, серотонина.
Информация, благодаря которой нейроны устанавливают только определенные связи с определенными нейронами, кодируется в структуре полисахаридных веточек мембранных гликопротеинов. Образование таких связей, не заложенных в период эмбрионального развития, является результатом опыта индивидуального организма и составляет материальную основу для хранения информации, определяющей особенности поведения данного организма.
Нейромедиаторы — это вещества, которые характеризуются следующими признаками. Накапливаются в пресинаптической структуре в достаточной концентрации. Освобождаются при передаче импульса. Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса. Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством. Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление — в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
Химическая классификация нейромедиаторов: Аминокислоты (и их производные) — К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
Таурин — образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин — это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты. Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин — Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора — Ацетил-КоА, синтезируется в нервных окончаниях. Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Норадреналин — медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Дофамин- медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.