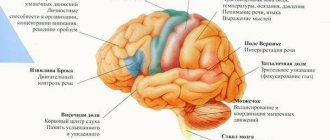
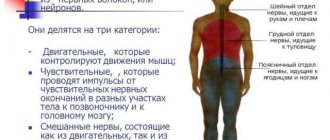
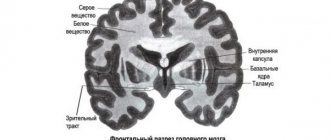
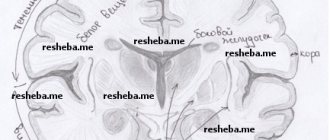
Projection zones of the cerebral cortex
Direct irritation of certain areas of the cerebral cortex leads to muscle spasms corresponding to the area of the cortex - the projection motor zone. When the upper third of the anterior central gyrus is irritated, a muscle spasm occurs in the leg, middle arm, lower face, and on the side opposite to the source of irritation in the hemisphere.
These seizures are called partial (Jacksonian). They were discovered by the English neurologist D.H. Jackson (1835-1911). In the projection motor zone of each cerebral hemisphere, all the muscles of the opposite half of the body are represented.
The cerebral cortex
,
substantia corticalis
; syn. cerebral cortex, cerebral cortex, mantle, mantle) - a layer of gray matter (1-5 mm thick) covering the cerebral hemispheres in mammals and humans; the highest department of the central nervous system, regulating and coordinating all the vital functions of the body during its interaction with the environment, K. b. n. - the material substrate of higher nervous and mental activity (although this activity is the result of the work of the entire brain as a single whole). In humans K. b. p. makes up on average 44% of the volume of the hemispheres, its surface is up to 1670 cm2.
There are ancient, old and new bark. The ancient and old cortex plays a significant role in the regulation of vegetative functions, the implementation of instinctive behavior, and in the need-emotional sphere. The functions of the neocortex are diverse and depend on the cytoarchitectonic zones. The neocortex (hereinafter referred to as the neocortex) plays an important role in cognitive processes, the organization of goal-directed behavior, and in humans in the implementation of higher mental functions.
There are projection cortical zones (see) - primary and secondary , and associative (see Associative areas
) -
tertiary and motor cortex . The main principle of the functional organization of projection
zones in the cortex is the principle of topical localization, which is based on clear anatomical connections between individual perceptive elements of the periphery and the cortical cells of the projection zones.
Projection sensory areas, including primary and secondary cortical fields
, receive and process information of a certain modality from the sense organs of the opposite half of the body (cortical ends of the analyzers according to I.P. Pavlov). These include the visual cortex located in the occipital lobe, the auditory cortex in the temporal lobe, and the somatosensory cortex in the parietal lobe.
Secondary projection zones also receive sensory signals predominantly from one modality; its neural organization creates conditions for the perception of more complex signal features.
Association cortical zones (tertiary) - make up 1/3 of the surface of the cerebral cortex in humans. Their role gradually increases among vertebrates up to humans. Having received maximum development in humans, A. k. z. They also adopted new, specifically human functions: speech, writing, intelligence, etc. A.k.z. developed in the anterior parts of the hemispheres, occupying most of the frontal lobes (prefrontal cortex) and at the junction of the projections of the main analyzers: visual, auditory and cutaneous-kinesthetic (posterior associative cortical zones). Nerve cells of A. k. z. react to stimuli of many modalities, and their responses arise not only to individual elements of an object, but also to its entire complexes.
Motor cortex
each hemisphere, occupying the posterior parts of the frontal lobe, controls and controls the motor actions of the opposite side of the body.
Functionally different areas of the cortex have a developed system of intra-cortical connections. The symmetrical cortical fields of both hemispheres are connected by fibers of the corpus callosum.
The system of intracortical connections and bilateral connections with underlying departments provide the possibility of forming functional systems that include structures of different levels.
The afferent and efferent projection zones of the cortex occupy a relatively small area. Most of the surface of the cortex is occupied by tertiary or interanalyzer zones, called associative zones.
Association areas of the cortex occupy a significant space between the frontal, occipital and temporal cortex (60-70% of the neocortex). They receive multimodal inputs from sensory areas. 52. Medial surface of the left hemisphere:
1 - precentral gyrus (motor zones); 2 - cingulate gyrus (part of the limbic system), responsible for visceral sensitivity; 3 - corpus callosum (main commissure); 4 - vault; 5 - frontal lobe; 6 - olfactory nerves, olfactory bulb and olfactory tract; 7 - temporal lobe; 8 - hippocampus (part of the limbic system); 9 - primary projection visual field (field 17); 10 - secondary projection visual field (field 18);
11 - occipital lobe; 12 - parietal lobe; 13 - posterior central gyrus (somatosensory areas)
thirds of the cortex and thalamic associative nuclei and have outputs to the motor areas of the cortex. Associative zones provide integration of sensory inputs and play a significant role in the processes of higher nervous and mental activity.
Associative areas
6>
All sensory projection areas and the motor cortex occupy less than 20% of the surface of the cerebral cortex.
The rest of the cortex constitutes the association region. Each associative area of the cortex is connected by powerful connections with several projection areas. It is believed that in associative areas the association of multisensory information occurs. As a result, complex elements of consciousness are formed.
In humans, like other mammals, the neocortex contains large areas that are not cortical centers of sensory or motor functions (nonspecific, or associative, areas), but their area significantly exceeds the area of the motor and sensory centers. Associative areas provide poorly developed connections between sensory and motor centers and, most importantly, are the morphological substrate of mental activity (consciousness, thinking, memory, emotions). This primarily applies to the frontal lobes. The frontal lobes also play a critical role in developing human behavior strategies. The associative cortex of the parietal and temporal lobes is involved in the formation of speech, in the perception and assessment of the location of one’s own body and its parts in space, as well as the three-dimensional spatial external world.
Association areas of the human brain are most pronounced in the frontal, parietal and temporal lobes.
Each projection area of the cortex is surrounded by association areas. Neurons in these areas are often multisensory and have greater learning abilities. Thus, in associative visual field 18, the number of neurons “learning” a conditioned reflex response to a signal is more than 60% of the number of background active neurons. For comparison: there are only 10-12% of such neurons in the projection field 17.
Damage to area 18 results in visual agnosia. The patient sees, walks around objects, but cannot name them.
The polysensory nature of neurons in the associative area of the cortex ensures their participation in the integration of sensory information and the interaction of sensory and motor areas of the cortex.
In the parietal associative area of the cortex, subjective ideas about the surrounding space and our body are formed. This becomes possible due to the comparison of somatosensory, proprioceptive and visual information.
Frontal associative fields have connections with the limbic part of the brain and are involved in organizing action programs during the implementation of complex motor behavioral acts.
The first most characteristic feature of the association areas of the cortex is multisensory
their neurons, and not primary, but rather processed information is received here, highlighting the biological significance of the signal. This allows you to formulate a program of targeted behavioral act.
The second feature of the associative area of the cortex is the ability to undergo plastic rearrangements.
depending on the significance of incoming sensory information.
The third feature of the associative area of the cortex is manifested in long-term storage
traces of sensory influences.
Destruction of the associative area of the cortex leads to severe impairments in learning and memory. The speech function is associated with both sensory and motor systems. Human speech and thinking are carried out with the participation of the entire cortex. At the same time, in the human cerebral cortex there are zones that are centers of a number of special functions related to speech. The nucleus of the motor analyzer of speech articulation, or the speech motor analyzer,
is located in the posterior sections of the inferior frontal gyrus (Broca's center, area 44, most often of the left hemisphere, was described by Dax (1835), and then by Broca) near the sections of the precentral gyrus, which are
analyzers of movements produced when the muscles of the head and neck contract.
Core of the auditory analyzer of oral speech
is closely connected with the cortical center of the auditory analyzer and is also located in the region of the superior temporal gyrus of the left hemisphere (field 22), in its posterior sections on the surface facing the lateral sulcus of the cerebral hemisphere.
Its function is to coordinate the auditory perception and understanding of the speech of another person and control its own speech. This center was described by Wernicke (1874). The motor and auditory speech centers are interconnected by a powerful bundle of axons. Speech functions associated with written speech - reading, writing
- are regulated by the angular gyrus of the visual cortex of the left hemisphere of the brain (field 39).
The core of the motor analyzer of voluntary movements associated with writing,
located in the posterior part of the middle frontal gyrus near the areas of the cortex of the precentral gyrus that control the movement of the hand and the combined rotation of the head and eyes in the opposite direction.
The speech motor analyzer analyzes the movements of all muscles involved in the act of articulate speech (pronunciation of words and sentences). In the center of the inferior frontal gyrus is the nucleus of the speech analyzer associated with singing.
In the middle third of the superior temporal gyrus there is the cortical end of the auditory analyzer, which belongs to the centers of the second signal system, which perceives the verbal designation of objects, actions, phenomena
, i.e. sensing signal signals.
Near the core of the visual analyzer is the core of the visual analyzer of written speech
, located in the angular gyrus of the inferior parietal lobule. Speech analyzers in right-handed people are localized only in the left hemisphere, and in left-handed people - only in the right.
When the motor center of speech is damaged , motor aphasia develops.
in this case, the patient understands speech, but cannot speak himself.
If the auditory center of speech is damaged, the patient can speak, express his thoughts orally, but does not understand someone else's speech, hearing is preserved, but the patient does not recognize words. This condition is called sensory auditory aphasia
.
The patient often speaks (logorrhea), but his speech is incorrect ( agrammatism
), and there is a replacement of syllables and words (
paraphasia
).
Damage to the visual center of speech leads to the inability to read and write. Isolated writing disorder - agraphia
, also occurs in case of dysfunction of the posterior parts of the second frontal gyrus of the left hemisphere.
In the temporal region located after 37, which is responsible for remembering words
. Patients with lesions in this field do not remember the names of objects. They resemble forgetful people who need to be prompted with the right words. The patient, having forgotten the name of an object, remembers its purpose and properties, so he describes their qualities for a long time, tells what they do with this object, but cannot name it.
The distribution of functions across brain regions is not absolute. It has been established that almost all areas of the brain have polysensory neurons, i.e. neurons that respond to various stimuli. For example, if field 17 of the visual area is damaged, its function can be performed by fields 18 and 19. In addition, different motor effects of irritation of the same motor point of the cortex are observed depending on the current motor activity.
If the operation of removing one of the cortical zones is carried out in early childhood, when the distribution of functions is not yet firmly fixed, the function of the lost area is almost completely restored, i.e. in the cortex there are manifestations of mechanisms of dynamic localization of functions that make it possible to compensate for functionally and anatomically disturbed structures.
Trace processes in the spinal cord after its irritation persist for a second; in the subcortical-stem regions (in the form of complex motor-coordinating acts, dominant attitudes, emotional states) last for hours; in the cerebral cortex, trace processes can be maintained according to the feedback principle throughout life. This property gives the cortex exceptional importance in the mechanisms of associative processing and storage of information, accumulation of a knowledge base.
The preservation of traces of excitation in the cortex is manifested in fluctuations in the level of its excitability; these cycles last 3-5 minutes in the motor cortex, 5-8 minutes in the visual cortex.
The main processes occurring in the cortex are realized in two states: excitation and inhibition. These states are always reciprocal. They arise, for example, within the motor analyzer, which is always observed during movements; they can also occur between different analyzers. The inhibitory influence of one analyzer on others ensures that attention is focused on one process.
The relationship between excitation and inhibition in the cortex manifests itself in the form of so-called lateral inhibition
.
With lateral inhibition,
a zone of inhibited neurons is formed around the excitation zone (simultaneous induction) and its length, as a rule, is twice as large as the excitation zone.
Lateral inhibition
provides contrast in perception, which in turn makes it possible to identify the perceived object.
In addition to the lateral
spatial inhibition, in cortical neurons after excitation, inhibition of activity always occurs and vice versa, after inhibition - excitation - the so-called sequential induction. In cases where inhibition is unable to restrain the excitatory process in a certain zone, irradiation of excitation occurs throughout the cortex. Irradiation can occur from neuron to neuron, along the systems of associative fibers of layer 1, and it has a very low speed - 0.5-2.0 m/s. In another case, irradiation of excitation is possible due to axon connections of pyramidal cells of the 3rd layer of the cortex between neighboring structures, including between different analyzers. Irradiation of excitation ensures the relationship between the states of the cortical systems during the organization of conditioned reflex and other forms of behavior.
Along with the irradiation of excitation, which occurs due to impulse transmission of activity, there is irradiation of the state of inhibition throughout the cortex. The mechanism of irradiation of inhibition is the transfer of neurons into an inhibitory state under the influence of impulses coming from excited areas of the cortex, for example, from symmetrical areas of the hemispheres.
6>
Date added: 2016-12-16; views: 2249; ORDER A WORK WRITING
Find out more:
3.10.2. Projection zones of the cortex
The use of methods for removing and irritating individual areas of the cortex made it possible to identify its motor (motosensory) areas. The most important motor area of the cortex is localized in the precentral gyrus
(fields
4
and b, according to Brodmann), lying anterior to the central (Rolandic) sulcus, which separates the frontal lobe from the parietal lobe (Fig. 3.34). Weak electrical
| Rice. 3.33 Cytoarchitectonic map of the human cerebral cortex according to Brodmann A—superolateral surface; B - medial surface. |
irritation of specific points in the precentral gyrus causes contraction of certain muscle groups on the opposite side of the body.
In the 30s. XX century W. Penfield, based on the results obtained during neurosurgical operations, established the presence of correct spatial projection
somatic muscles of various parts of the body to the motor cortex.
Having discovered the somatotopic organization of the motor cortex, W. Penfield noted that the areas that control the movements of the hands and facial muscles are the most extensive and are characterized by the lowest threshold. Subsequently, another motor area was discovered on the medial surface of the hemisphere next to this primary motor area and, thus, multiple representations of peripheral formations in several motor areas of the cortex were shown. Due to the fact that these areas, in addition to the motor output from the cortex, have independent sensory inputs from skin and muscle receptors, they were called the primary
and
secondary motosensory cortex
(McI and McII).
| Rice. 3.34 Simplified map of the division of the lateral cerebral cortex into functional fields according to Kleist The numbers indicate Brodmann's cytoarchitectonic fields (see Fig. 3.33). |
Subsequent experimental studies showed that in the postcentral gyrus
(fields 1,
2, 3)
is
the first somatosensory (sensorimotor) area,
where afferent projections from skin receptors and the motor system come through specific nuclei of the thalamus (see Fig. 3.33). As in the motor cortex, these projections have a somatotopic organization, with points close to each other on the surface of the body having representation in neighboring areas of the postcentral gyrus. Removal of certain zones of the postcentral gyrus leads to the loss of discriminatory sensitivity in certain parts of the body. Electrical stimulation of the postcentral gyrus in patients undergoing neurosurgical operations (under local anesthesia) causes a feeling of numbness or tingling in certain areas of the body surface.
a second somatosensory area has been discovered in carnivores and primates
localized ventral in the area
of the lateral
(Sylvian)
fissure,
which separates the temporal lobe from the parietal and frontal.
There is also a somatotopic projection of the body surface here, but less clear than in the first somatosensory area. It turned out that the first and second somatosensory areas, in addition to afferent inputs, contain motor outputs and, therefore, it is more correct to call them primary
and
secondary sensorimotor zones
(Sm I and Sm II).
Therefore, we can judge the presence of four sensitive areas in the cortex
(Sm I, Sm II, McI and McII), arranged here in descending order of their importance. Likewise, there are four motor areas in the cortex (McI, McII, CmI, CmII), and the leading role in this series belongs to the primary motosensory cortex.
Among other projection zones of the cortex, the primary visual area
(field
17),
localized in
the occipital lobe.
In the projections of retinal receptors onto area
17,
as well as in other sensory systems, a precise topographic order is revealed, with the same halves of the retinas projecting into each hemisphere. The combination of the visual fields of both eyes that occurs in each hemisphere underlies binocular vision. Damage to a limited area of the visual cortex can lead to partial blindness, which results in loss of a small portion of the visual field. Electrical stimulation of the visual cortex can produce visual sensations.
Next to the primary visual area is the secondary one,
occupying fields
18 and 19.
These fields are related to functions such as visual attention and control of eye movements (see section 4.8.4).
Currently, the primary projection zone of the auditory analyzer has been well studied,
which is localized in the upper edge
of the temporal lobe
(fields
41
and
42).
For this area, as well as for other projection zones, a tonotopic representation of the spiral (Corti) organ of the cochlea has been described, consisting in the fact that different parts of the organ are projected to certain loci of the auditory cortex (see section 4.5.2).
Damage to fields 41
and
42
makes it difficult to perceive speech, spatially localize a sound source, and recognize the temporal characteristics of sound.
Within each lobe of the cerebral cortex (Fig. 3.35), next to the projection zones there are fields that are not associated with the performance of any specific sensory or motor function. Such fields constitute the association cortex,
for which neurons tend to respond to stimulation of various modalities and thus participate in the integration of sensory information and in providing connections between the sensitive and motor areas of the cortex.
These mechanisms are the physiological basis of higher mental functions.
The association cortex includes a number of areas of the parietal, temporal and frontal lobes. Unfortunately, the boundaries of these areas and the cortico-cortical connections between them have not been fully identified, and most physiological
| Rice. 3.35 The four main lobes of the cerebral cortex (frontal, temporal, parietal and occipital). Side view 1 - prefrontal associative cortex, 2 - premotor cortex, 3 - primary motor cortex, 4 - central sulcus, 5 - primary somatosensory cortex, 6 - parietal lobe, 7 - secondary visual cortex, 8 - occipital lobe, 9 - primary visual cortex, 10 - parietal-temporo-occipital associative cortex, 11 - primary auditory cortex, 12 - secondary auditory cortex, 13 - temporal lobe, 14 - limbic associative cortex, 15 - lateral sulcus, 16 - frontal lobe. |
data is based on symptoms of damage to associative areas in animals and on clinical observation data.
At the same time, it has been established that the parietal associative fields,
which in the process of evolution are built over the visual projection zone, participate in the assessment of biologically significant information and in the perception of spatial relations of the surrounding world.
Frontal lobes
(fields
9-14),
which have extensive bilateral connections with the limbic system of the brain, control
the assessment of behavioral motivation
and
the programming of complex behavioral acts.
The involvement of the frontal lobes in
movement control has been established.
For example, damage to the frontal lobes causes patients to tend to repeat motor acts without apparent correspondence with external circumstances.
Experiments on monkeys have shown that damage to the frontal lobes impairs memory for the location of objects, and this process is associated with dopaminergic structures that are rich in the frontal lobes.
Apparently, the frontal lobes, along with the parietal lobes, are involved in the integration of information about space and time.
Brain structure and starting settings
Knowing how a neuron works is not enough to understand what causes rational and intelligent behavior.
Evolution has performed an amazing feat of creating a system from relatively simple elements that can amaze with incredible success in interacting with its environment. It is not enough to take a certain mass of connected neurons (even folded in layers), connect sensors and outputs to it and get at least some semblance of a brain. The main field of work of evolution over millions of years is not the neuron, but the structure and internal organization of nerve cells in the nervous system. In the previous part we talked about the structure of the cerebellar cortex and its example shows that structure and organization is fundamental to its functions. Let's look at how the cerebral cortex is organized, the structure thanks to which Man has become the most successful species on Earth. Table of contents
1. Nervous system simulator. Part 1. Simple adder 2. Nervous system simulator. Part 2. Modulated neuroelement 3. Nervous system simulator. Part 3. The associative neuroelement 4. Memory, memory consolidation and granny neurons 5. Simulation of emotions or the electronic sense of novelty 6. The amazing cerebellum 7. Brain structure and starting settings
It is known that the human cerebral cortex consists of six conventional layers, but it came to this form over many stages of evolution. Therefore, to begin with, we will consider a simplified version with 2 - 3 layers, because this option also occurs in nature. It is more advantageous to study any biological structure from the perspective of its evolutionary development. Evolution followed two paths of development of the cortex: an increase in the number of layers and an increase in the total area of the cortex.
The second feature of the cortex is the presence of so-called cortical columns. We can say that the cortical column is the logical unit of structure of the cortex. The growth of the cortex itself during embryonic development is carried out by division, self-copying of entire columns. And this is logical if we want to maintain some structure while growing.
The cortical columns are based on pyramidal neurons. Pyramidal neurons are one of the largest neurons of the nervous system, and therefore the most studied, since their size allows them to be influenced using special electrodes without destroying the cell. These neurons have a developed system of dendrites, which extends throughout the column, as well as an axon, which in some cases is the source of incoming or outgoing signal. Despite the fact that there may be several such neurons in the first layers of the cortical column, they all work as one neuroelement. The activity of one pyramidal cell often means the activity of the entire group of cells. The evolutionary development of the nervous system was previously focused on reliability, and duplication or distribution of one functionality among groups of cells is a ubiquitous phenomenon. It is impossible to imagine a scheme created by nature in which the death of just one cell would lead to disruption of the functioning of an entire functional unit. We can say that the activity of a group of pyramidal cells in a column indicates the activity of the column itself.
Speakers can be divided into two types: signal generating and signal receiving. The type of column depends on what kind of axons the pyramidal neurons have: afferent or efferent. If there are afferent axons, that is, those bringing a signal, then the column will be activated when a signal is received; the signal may be relayed if there is an efferent axon. In the presence of efferent axons, the result of column activity will be the formation of an outgoing signal.
The dendrites of pyramidal neurons extend to the upper layers of the cortex, where they extend beyond the boundaries of their cortical column, thus causing local interaction between neighboring columns. One of the forms of local interaction is lateral (side) inhibition. Inhibition of neighboring columns is carried out through special inhibitory neurons that are part of the cortical columns. Mainly inhibitory neurons exert their effect on pyramidal neurons, preventing their activation.
Lateral inhibition is exerted on adjacent surrounding columns. It allows you to make the boundaries of the active column areas clearer and the activity areas more localized. Thanks to lateral inhibition, an obstacle to the strong spread of excitation arises.
In addition to lateral inhibition, lateral drive is also present. By adjusting the balance between these two factors, it is possible to very finely regulate the level of overall activity in the brain. For example, sleep requires a reduction in the level of activity; this requires strengthening lateral inhibition and weakening drive. This is done through special chemical signals and mediators.
The effect on neighboring columns does not always take the form of a uniform concentric distribution. The entorhinal cortex is characterized by a distribution that allows arousal to spread more easily in a triangular grid. These are the so-called grid cells, which help animals navigate in space by additionally modifying the image of their place of stay.
The next element of the cortical column is a layer of many relatively small stellate neurons. This layer is usually called granular. The neurons of this layer, due to their size, are less studied than pyramidal neurons. It is the neurons of this layer that can be assigned the main role in the computational processes of the brain; thanks to their work, the formation of associative connections and the formation of images occurs. “Calculations” occur according to the principles of mutual attraction of excitation. Neurons of the granular layer are an associative neuroelement. And the pyramidal cells are assigned the role of a simple adder; these elements seem to separate and highlight images. Pyramidal neurons will be activated when a significant portion of the neurons in the granular layer of the column are excited.
Connections between stellate neurons can freely extend beyond the boundaries of their cortical columns; we can say that the granular layer is almost continuous.
Memory, memory consolidation and granny neurons
Formation of images
Separation of an image by a layer of simple adders
This version of the cortical column and cortical structure is very simple and may even seem primitive, but when used on a large scale and with the right setup, it can produce a structure with high computational performance. Nature has always chosen the simplest, most reliable and effective solutions, and our nervous system is no exception to this rule. I often come across the opinion that a neuron is like some kind of supercomputer or quantum computer that makes complex calculations using some kind of vibrations of ions on its membrane or quantum mechanisms. Even the summation algorithm in the neurons of a conventional perceptron is more complex than in its biological counterpart; in a perceptron, it matters which synapse the signals come from, but in a biological one, only the total amount of impact is important. Stereotypical thinking about the unattainable complexity of the brain can hinder understanding the nature of a biological system.
Surprisingly, the described structure is universal for various types of cortical areas: sensory, motor and associative. There may be variations in the relative thickness of the different layers depending on the function of the cortical area. For example, the motor cortex has an increased layer of pyramidal neurons relative to the granular layer, because the signals from the motor cortex must be clear and strong. And associative areas are characterized by an increased layer of stellate neurons to provide the greatest flexibility in the formation of associative reflex arcs.
Cortical areas are intertwined with many connections, this is done thanks to axons, long processes of neurons. Bundles of axons form nerves, which are the so-called white matter. These nerves can connect both adjacent areas and areas of opposite halves of the brain. Moreover, the architecture of these connections is determined by the evolutionary development of the brain and partially acquired experience and learning, but for different people the picture of these connections will be similar. There are several scientific projects related to mapping these connections, for example, the Human Connectom Project.
Let's look at the principles by which these communications are organized.
The presented diagram is only an example for understanding the principles of the organization. Real circuits in the biological nervous system are many times more complex.
Let's imagine a certain receptor field with a number of receptors of the same type that allow us to obtain certain information about the environment. Certain receptor fields form combined signals, for example, the retina of the eye. Such signals require certain analysis capabilities. Representative columns of these receptors will be distributed throughout the sensory cortex with a certain density while maintaining the topology of the location of the receptors in the receptor field (A). According to the principles of mutual attraction of excitation, certain areas of excitation will form on the cortex, which will be the image of the received combined signal. The primary sensory cortex usually has the highest neuroplasticity, i.e. any combination of excited columns will be processed without taking into account previously received information. The resulting image will be read by other columns, also distributed with a certain density throughout the sensory cortex. These columns will transmit information for further processing to the following areas of the cortex. The nature and density of the “reading” columns provide a certain filter for the resulting images. It is not difficult to understand that this method of processing leads to a significant loss of information received from the receptors; the resulting images do not provide unambiguous information about which receptors were activated. Evolution has chosen two ways to eliminate this problem. Firstly, there is an excess number of receptors, which compensate for the drop in information content after processing. Secondly, if you duplicate information from receptors in another area, but with a violation of the topology of the location of the representative columns in relation to the location of the receptors in the receptor field (B), i.e. Let's confuse them. Then, with many different combinations of receptor activity in two areas, different combinations of images will be formed, which gives more information content and more features that we can identify. Naturally, it should be understood that the “confusion” of signals occurs in a strictly defined way; for example, if you “confuse” the entire field of the retina, this will not give any benefit. Entanglement occurs in small fragments of receptor fields. And of course we can, if necessary, duplicate and confuse the signals more than once. In the nervous system, an example of this division is the dorsal and ventral visual processing pathways.
The general principle of information processing by the cerebral cortex is the sequential transmission of information from area to area with a decrease in the density of connections. Moreover, with each subsequent level, the neuroplasticity of the areas decreases, which adds memory and experience in processing previous information to the calculations. Thus, from the processed information, some key features can be identified that will be associated with specific nerve cells. Simply put, in areas of the cortex with a higher level of processing, the form of excitation distribution that corresponds to the most frequent form of the processed signal will be formed.
The formed image “abc” by a combination of columns a, b and c with frequent repetition or emotional reinforcement will be executed in the future even if the activated combination is incomplete and noisy.
At each level of information processing, a certain time is spent; if it is necessary to analyze the dynamics of changes in information over time, then it is possible to duplicate part of the information from each level into one area. An analogue in biology is the visual information processing area MT (V5), the middle temporal cortex, which collects information from areas V1, V2, V3..., this area is responsible for the perception of movement. When this area is damaged, akinetopsia occurs - the inability to perceive movement.
Associative areas are arranged relatively simply; representations of various sensory and motor areas are located here. Moreover, for motor representations, the presence of bilateral connections is important, either these are axons that can work in both directions, or they are adjacent and jointly working columns at both ends, or two axons of the same column working in different directions. There should be many such areas with different patterns of arrangement of representations, so that the possibility of forming reflex arcs for various combinations is equally possible. Conditioned reflexes are formed in such areas, so these areas should have reduced plasticity.
All connections in the motor and primormotor areas must be bilateral in nature; this is necessary for the formation of associative connections. Essentially, the primormotor area must form reflex arcs of sequences, from one focus of excitation to another. In order for there to be more variability, representations are needed, leading to actions to be repeatedly mixed, this is similar to the situation with sensory processing only in reverse order. Also, all motor actions interact with a specially designed area that coordinates actions in time - the cerebellum.
The presented diagrams are a strong simplification of how it is wired in the brain, and the creation of logical structures similar to the human nervous system is impossible without the participation of specialists in the field of neurobiology and scientists studying the connectome.
What about the other layers?!
– Indeed, I only talked about three layers of the cortex, but in the human brain there are six layers in the cerebral cortex. The cerebral cortex turned out to be a fairly successful product of evolution, even with a small number of layers. The principle of evolution: don’t touch what works. Therefore, all new layers in the cortex are an addition to existing layers. If you look at the layers of the human brain, you can see that we do not have six visible layers, but two logical layers, the structure of which is similar and repeats. Evolution simply repeated existing structures in order to increase productivity.
The pyramidal cells of the outer layer are smaller than the pyramidal cells of the first layer, which means they generally have a higher threshold of sensitivity to activating factors.
The granular layers will operate under similar conditions, but presumably the stellate neurons in the outer layer have lower plasticity, meaning that under some conditions the patterns of activity in the granular layers may differ, despite the fact that the input signals are the same. Thanks to these two logical layers, two modes of activity of the cortical column arise. First: full activity mode, when the pyramidal cells of both layers are activated, the entire column is activated. Second: partial activity mode, when only the upper additional layer is activated. These two operating modes of the speaker can be compared to a person’s ability to speak in a full voice and in a whisper; a whisper is partial activity, and a full voice is full activity.
What does this give? For the sensory cortex, this is an additional level of information processing, as well as the ability to work with images of these areas without activation through receptors. In other words, it gives you the opportunity to work with your imagination. For associative areas, this is an additional level of abstraction, the formation of associations between images that have fewer common features, since the sensitivity threshold of the pyramidal cells of the additional layer is higher. For the motor and primortial cortex, this is an opportunity to work out some movements without directly performing them. Only when the column is fully activated do actions occur; actions with partial activation remain in our imagination.
Of course, there are areas in the brain that control the operation of speaker modes, just as we can very easily change the nature of our speaking from a whisper to full volume. If you increase the level of inhibition in the column, then it is possible that it will only be partially activated; if, on the contrary, you disinhibit the column, then some thoughts can immediately be translated into actions.
Fantasy and a high level of abstract thinking have made humans the most successful species on Earth.
Even if we correctly configure the areas and connections between them, this will not be enough to obtain a working model. Unconditioned reflexes are required. A person is born with a rich set of reflex mechanisms, carefully selected by evolution.
Setting up unconditioned reflexes for the model is an important point, given the fact that learning new reflexes always occurs on the basis of existing reflexes. If any action is not involved in some unconditioned reflex, then it will be impossible to learn to control this action. Biological systems do not initially have “clear” reflexes. After birth, we cannot precisely control our limbs or, for example, walk. This is due to the fact that it is impossible to determine in advance some body parameters, the size of the limbs, their weight, the force created by the muscles, etc. Moreover, these parameters also change dynamically during the growth of the organism. Therefore, many unconditioned reflexes have a certain field of action in the response, and in the head a field of receptors that activate it. An emotional mechanism associated with unconditioned reflexes is also laid down, which will trigger reflexes for correction at a certain point in development.
Let's consider the mechanism for adjusting reflexes using the example of baby babble. In accordance with a certain stage of development, the babbling mechanism is launched, i.e. there is an almost spontaneous launch of “fuzzy” reflexes. When they are started, the child begins to utter various sounds, sometimes these same reflexes are provoked by a sound heard from the outside. The sounds produced often do not correspond to what is expected, i.e. do not correspond to the sounds that trigger the reflex, or trigger hypothetically. The child hears the sounds he himself makes, receiving feedback between the command and the resulting action. Next, the emotional mechanism of novelty comes into force, which connects the center of the need for novelty with a speech act, which gives a new association between the heard sound and the internal motive that prompted the action. Which leads to repeated repetition of the action leading to a saturation of the feeling of novelty. It is claimed that during the period of babbling a child pronounces all the sounds of all languages on Earth. Repeated repetition of sounds leads to the formation of clear patterns of action in accordance with the desired result.
Mastery of the motor-motor system occurs in a similar way. Initially, infants' movements are almost chaotic; there is only an increase in motor activity as a reaction to an emotional stimulus. But over time, a comparison of movements and visual perception, tactile and perception of body position occurs.
Some unconditioned reflexes are not so primitive; in certain cases, template images are embedded in the nervous system, and it is almost impossible to transfer such templates from biological systems to a computer model. Humans have an innate ability to recognize the emotions and movements of individuals of their own species. Therefore, there will be a need to implement some workarounds for some aspects of training.
To bypass the process of long-term training of the motor-motor system through numerous attempts at crawling, standing up, walking and a series of falls, it is possible for android robots to use the method of intercepting control.
A person can transfer his experience of body control to a robot through special devices and motion interception technologies. In a model of the nervous system of a robot with intercepted control, during motor actions the corresponding representations will be activated, as if these movements were performed by the robot itself. Thanks to this, the necessary images and associative connections would be formed. For example, during training with the command: “Raise your hands” - the teacher in the movement interception mode raises his hands, this would lead to the formation of a conditioned reflex between the command and the action, and an associative connection would be formed between the command and the image formed by the processing of position sensors bodies.
In the process of training an electronic model of the brain, it is always possible to control the plasticity of the desired areas, and it is also possible to look “inside” the learning process and highlight, label and strengthen the resulting images. This should significantly speed up the learning process of artificial nervous systems in relation to human learning. As has already become clear, a model built according to the principles described will learn for the most part like a person, without separating learning and interaction with the environment.
Instead of a conclusion
This series of articles ends the next stage of research and development.
The program developed for modeling, although limited in its functionality, made it possible to form a theoretical basis for further work. Of course, the resulting theory still requires further development, as well as practical confirmation and testing. But now it will allow us to take a different look at the problem of modeling the nervous system. The next step is the development of a new modeling environment that will allow us to create models of larger-scale processes occurring in the nervous system. And also to implement a number of ideas, observations and theoretical conclusions. I am not a scientist, and my main activity is not related to the development of intelligent systems, nor is it related to neurobiology.
But I will make every effort to ensure that the project comes to its logical conclusion. I will be glad to consider your suggestions, advice and recommendations, as well as constructive criticism. I will definitely talk about the development news of the project on the pages of GeekTimes and my YouTube channel. Thanks to all! PS It would be inhumane to post the sources without explanation. Therefore, I will post a link to the source and some explanations on the algorithm and logic in a separate article.