In the spinal cord, the reflex rings of the skeletal muscles are interconnected with the reflex rings of the internal organs. Therefore, when receptors are stimulated, afferent nerve impulses also cause excitation of efferent neurons innervating the heart, blood vessels, glands and other organs. At the same time, hormones and other chemical products enter the blood and the central nervous system becomes the object of their influence.
Consequently, the spinal cord contains neurons that cause somatic (motor) and autonomic reflexes.
The basic properties of the nerve centers of the spinal cord are of subordinate importance, since the leading role in unifying and regulating all the activities of the body in its interaction with the outside world belongs to the higher department of the nervous system - the cerebral hemispheres.
Determination of excitability of spinal cord centers
The excitability of the spinal cord is determined by the duration of the latent period of the reflex and the magnitude of the reflex reaction. The latent period of the reflex is much longer than when irritating a neuromuscular drug. It consists of the time of excitation along the afferent pathway, through the central nervous system and along the efferent pathway. Detailed studies also take into account the latency period of the receptor and effector.
The time required for excitation to pass through the central nervous system or required for the transition of excitation from afferent to efferent neurons is called central reflex time , or reduced reflex time . Its duration depends on the number of neurons and synapses involved in the implementation of this reflex. The longer it is, the more neurons and synapses there are along the path of excitation through the central nervous system, i.e., in the reflex ring, since the conduction of excitation is delayed in each neuron and especially in each synapse.
The central reflex time for a two-neuron reflex “arc”, in which there is only one synapse, is the smallest. The duration of the excitation delay in each synapse of the central nervous system (or in the myoneural apparatus) is minimally equal to 0.3 ms (in each on average no more than 0.5 ms).
The tendon reflex time is the shortest; for example, knee - 5.3 ms, of which 3.1 ms accounts for the conduction of excitation along peripheral pathways, and 2.2 ms for central time. When the stimulation is repeated, the central time can decrease to 1 or even 0.3 ms. The duration of the central reflex time is greatest with weak stimuli. With complex somatic reflexes, the central reflex time increases to 10-20 and even 500 ms, and with vegetative reflexes - up to tens of seconds.
Changes in excitability
. Immediately after each nerve impulse, repeated stimulation causes absolute and relative refractory phases in the neurons of the spinal cord, and then the exaltation phase begins. These phases are longer than in nerve fibers.
After the exaltation phase, excitability decreases. The rhythm and strength of the reflex reaction depend on changes in the excitability and lability of neurons. Due to the occurrence of excitation in some neurons of the spinal cord, their excitability increases, and due to the occurrence of inhibition in other neurons of the nervous system, the excitability of these neurons decreases. Consequently, the excitability of spinal cord neurons constantly changes due to changes in excitation and inhibition, and therefore there is no constant threshold of reflex stimulation.
Excitability changes depending on the influx of impulses coming from the receptors. Synapses for many days can be in a state of sharply reduced excitability or, conversely, in a state of increased excitability, which in the first case is accompanied by the absence of motor reflexes, and in the second - an increase in reflex movements.
Test with answers: “Structure and functions of the spinal cord”
1. With a damaged spinal cord at the level of the cervical spine, it is possible to turn the head, talk, make chewing movements, is this true: a) yes + b) no c) partly
2. In a spinal frog (without a brain), motor reflexes are preserved, is this true: a) no b) yes + c) not studied
3. The spinal cord performs a conductive function, is this true: a) no b) not studied c) yes +
4. When the spinal cord is damaged at the level of the cervical spine, any movements become impossible, is this true: a) no + b) yes c) partly
5. The spinal cord consists of... segment: a) 28 b) 42 c) 31 +
6. The spinal cord consists of 42 segments, is this true: a) yes b) no + c) partly
7. The thickness of the spinal cord is about 4 cm, is this true: a) yes b) different c) no +
8. The spinal cord performs a reflex function, is this true: a) no + b) yes c) partly
9. Loss of sensitivity in certain areas of the body is observed with: a) damage to the anterior horns of the spinal cord b) damage to the anterior roots of the spinal nerves c) damage to the dorsal roots of the spinal nerves +
10. Paralysis of a certain group of muscles is observed when: a) the spinal cord is destroyed b) damage to the dorsal roots of the spinal nerves + c) damage to the anterior roots of the spinal nerves
11. From the spinal cord, excitation passes through: a) the anterior roots of the spinal nerves + b) mixed nerves to the brain c) the dorsal roots of the spinal nerves
12. Excitation passes to the spinal cord along: a) anterior roots of the spinal nerves b) dorsal roots of the spinal nerves + c) mixed nerves from the brain
13. Interneurons are located: a) in the anterior horns of the gray matter of the spinal cord b) in the anterior roots of the spinal nerves c) in the dorsal horns of the gray matter of the spinal cord +
14. Motor neurons are located in: a) nodes of the dorsal roots of the spinal nerves b) dorsal horns of the gray matter of the spinal cord c) anterior horns of the gray matter of the spinal cord +
15. Sensory neurons are located in: a) ganglia of the dorsal roots of the spinal nerves + b) anterior roots of the spinal nerves c) dorsal horns of the gray matter of the spinal cord
16. The white matter of the spinal cord is formed by the processes of neurons, is this true: a) no b) yes + c) not studied
17. Nerves and blood vessels pass through the spinal canal, is this true: a) yes b) not studied c) no +
18. The gray matter of the spinal cord is formed by the processes of neurons, is this true: a) yes b) no + c) not studied
19. In which section is thickening of the spinal cord observed: a) sacral b) thoracic c) cervical +
20. In which section is thickening of the spinal cord observed: a) thoracic b) lumbar + c) sacral
21. Which reaction (action) of the body with the help of a reflex arc is directly related to the spinal cord: a) sneezing + b) eye movement c) squats
22. What are the ascending pathways: a) transmission of signals from the brain to the spinal cord b) transmission of signals from the spinal cord to the brain + c) transmission of signals from the brain to the organs
23. What is gray matter: a) accumulation of nerve cell processes b) accumulation of myelin fibers c) accumulation of nerve cells +
24. What fills the space between the spinal canal and the spinal cord: a) white matter b) adipose tissue + c) gray matter
25. What fills the space between the spinal canal and the spinal cord: a) gray matter b) adipose tissue c) blood vessels +
26. What fills the space between the spinal canal and the spinal cord: a) white matter b) cerebrospinal fluid + c) adipose tissue
27. In what sequence are the membranes of the spinal cord located (from the center to the periphery): a) soft, arachnoid, hard + b) hard, arachnoid, soft c) hard, soft, arachnoid
28. What is a spinal filum: a) nerve fibers extending from neurons b) a hollow canal passing through the center of the spinal cord c) a narrow process of the spinal cord fused with the periosteum of the coccyx +
29. Where does the spinal cord end: a) at the level of the first lumbar vertebra + b) at the level of the first sacral vertebra c) at the last lumbar vertebra
30. Which part of the brain is the spinal cord connected to: a) forebrain b) medulla oblongata + c) diencephalon
Coordinating mechanisms of the spinal cord
Coherence, subordination of contractions of individual muscles or their groups, based on the interaction of excitation and inhibition in the nervous system, is referred to as coordination.
The stimulation of a group of neurons is accompanied by a change in the functional state of neighboring neurons and their inhibition. According to us, each reflex is the result of the interaction of excitation and inhibition in the central nervous system, the coordinated and subordinate activity of many neurons. It is thanks to the coordination of neurons, which arose in the process of phylo- and ontogenetic development, that subtle movements and isolated motor reflexes can be carried out instead of the general mass reflex movements inherent in organisms in which the nervous system is at the lower stages of phylo- and ontogenesis. Coordination characterizes not only complex, but also the simplest reflexes.
Participation of the spinal cord in the regulation of muscle tone
Regulation of muscle tone is of a reflex nature and is carried out through myotatic and postural-tonic reflexes.
The myotatic reflex is a stretch reflex that begins with irritation of the muscle spindle. Almost every muscle contains muscle stretch receptors, called “muscle spindles” because of their shape (Fig. 2).
The muscle receptor consists of striated intrafusal muscle fibers surrounded by a connective tissue capsule and has the shape of a spindle. The length of intrafusal muscle fibers is 4 - 7 mm, thickness 15 - 30 µm. Muscle spindles are located parallel to the skeletal muscle and their ends are attached to the connective tissue membrane of the bundle of extrafusal muscle fibers using tendon-like strips of connective tissue 0.5 - 1 mm long.
The mechanism for regulating muscle tone is presented in Figure 2. Stretching of extrafusal muscle fibers (with relaxation of striated muscle fibers) leads to stretching of the muscle spindle. When the muscle spindle is stretched, the nuclear bursa is stretched and the stretch receptor is excited. Impulses enter the spinal cord to α-motoneurons. Excitation along the axons of α-motoneurons causes contraction of extrafusal muscle fibers.
Fig.2. Scheme of regulation of muscle tone.
1 – extrafusal fibers; 2 – muscle spindle; 3 – intrafusal
fibers; 4 – efferent nerve fibers; 5 – alpha motor neurons;
6 – gamma motor neurons
Gamma loop:
A number of effects from muscle and tendon receptors go not only to α-motoneurons, but also along the axon collateral to γ-motoneurons: γ-motoneurons are excited → send impulses to intrafusal fibers → intrafusal fibers contract → muscle spindle stretches → receptor is excited → impulses go to spinal cord → activation of α-motoneurons → contraction of extrafusal fibers.
Cervical posture-tonic reflexes
spinal cord are aimed at maintaining posture. With their help, muscle tone is regulated. These reflexes arise from the proprioceptors of the muscles and fascia of the neck. Switching from these receptors occurs at the level of the cervical spinal cord and leads to changes in muscle tone when the position of the head and neck changes.
Segmental and intersegmental principle
Spinal cord works
The spinal cord consists of 31-32 segments. Each segment corresponds to two pairs of roots (corresponding to the number of segments, 31-32 pairs of anterior motor roots exit the spinal cord and 31-32 pairs of posterior sensory roots enter it). When projecting segments of the spinal cord onto the vertebrae, it is necessary to take into account the discrepancy between the lengths of the spinal cord and the spine. In the cervical region, the segments are located 1 vertebra higher than the corresponding vertebra; upper thoracic - by 2, lower thoracic - by 3 (example: the V cervical segment is located at the level of the IV cervical vertebra, the V thoracic - at the level of the III thoracic vertebra, the XI thoracic - at the level of the VIII thoracic vertebra, etc.). The lumbar and sacral segments and vertebrae are in great disparity: the lumbar segments are at the level of the X, XI, and XII thoracic vertebrae; sacral – XII thoracic and I lumbar. There are no morphological boundaries between the segments of the spinal cord, therefore the division into segments is functional and is determined by the distribution zone of the dorsal root fibers in the segment and the zone of cells that form the anterior root. The part of the body innervated by the fibers of one segment is called the metamer (the area of the skin innervated by the sensory fibers of the segment is called the dermatome). Each segment innervates through its own metamer. This is the segmental principle of the spinal cord. The intersegmental principle of the spinal cord is that each segment of the spinal cord innervates not only its own metamer, but also the metameres of the upper and lower adjacent segments. As a result of the overlap of nerve fibers, each metamer of the body is innervated by three segments and transmits signals to 3 segments of the spinal cord, which increases the reliability of regulatory mechanisms.
Structural organization and functions of the medulla oblongata
The medulla oblongata is a continuation of the spinal cord and is about 25 mm long. Unlike the spinal cord, its gray matter is located not in the center, but on the periphery in the form of nuclei.
In the medulla oblongata there are nuclei of the VIII-XII pairs of cranial nerves through which the regulation of sensitive (sensory), motor (somatic) and autonomic (parasympathetic) functions is carried out: sensory nuclei of the vestibular-cochlear nerve (n.vestibulocochlearis, VIII), nuclei of the glossopharyngeal nerve (n. glossopharyngeus, IX), nuclei of the vagus nerve (n. vagus, X), motor nucleus of the accessory nerve (n. accessorius, XI), motor nucleus of the hypoglossal nerve (n. hypoglossus, XII). In addition, in the RF of the medulla oblongata there are vestibular nuclei (medial - Schwalbe's nucleus, superior - Bekhterev's nucleus, lateral - Deiters' nucleus and inferior - Roller's nucleus), which receive impulses from the receptors of the semicircular canals.
Functions of the medulla oblongata
The medulla oblongata performs sensory, conductive and reflex functions.
— Touch functions
are carried out due to the receipt of afferent information into the medulla oblongata from taste receptors, auditory and vestibular receptors along sensory fibers to the nuclei of the corresponding cranial nerves and from receptors of the thoracic and abdominal organs (see above). At the level of the medulla oblongata, a primary analysis of the strength and quality of the stimulus is performed.
— Conductor functions
. All ascending and descending tracts of the spinal cord pass through the medulla oblongata. Here are the intersections of the descending pyramidal paths and the ascending tract of Gaulle and Burdach.
— Reflex functions
.
The centers of numerous reflexes are localized in the medulla oblongata. For example, a number of protective reflexes: gag reflex
(from the receptors of the root of the tongue, pharynx, stomach, intestines, peritoneum, vestibular apparatus → along the fibers of the glossopharyngeal, vagus or vestibular nerve → to the vomiting center of the medulla oblongata → along the efferent fiber of the vagus nerve to the esophagus, stomach, intestines and through the spinal motor centers to the diaphragm and abdominal wall muscles);
cough reflex
(from the receptors of the larynx, trachea and bronchi → along the sensory fibers of the vagus nerve → to the cough center of the medulla oblongata → efferent impulses to the spinal centers of the respiratory muscles);
sneezing reflex
(from the receptors of the nasal cavity → to the sneezing center of the medulla oblongata → efferent impulses to the spinal centers of the respiratory muscles), as well as
reflexes of lacrimation and eyelid closure.
Also, the medulla oblongata, together with other structures of the brain stem, is involved in the implementation of
postural maintenance reflexes (vestibular postural-tonic reflexes).
In the medulla oblongata, in addition to the centers of protective reflexes, in the area of the reticular formation there are vital centers - the vasomotor and respiratory centers.
Features of the structural organization of the reticular formation
brain stem
The reticular formation (RF) is located in the gray matter of the medulla oblongata, midbrain, diencephalon, and partly the spinal cord and regulates the level of activity of the cerebral cortex, cerebellum, thalamus, and spinal cord. The RF is formed by neurons that have long, poorly branched dendrites and a short, well-branched axon. The morphological structure of the reticular formation is very reminiscent of a mesh, and it is on this basis that the German anatomist Otto Deiters called this structure “reticular formation” (Latin reticulum - mesh, formatio - formation) (1865). RF neurons receive information from different receptors. These are multimodal neurons that have large receptive fields, have spontaneous activity, increased excitability, high lability (up to 1000 pulses/sec), and high sensitivity to metabolites, hormones, barbiturates and other pharmacological drugs.
Features of excitation irradiation in the spinal cord
Excitation entering the spinal cord via an afferent neuron spreads to the above and below located segments. This process of spreading excitation is called irradiation.
The irradiation of excitation occurs especially clearly due to an increase in the strength of stimulation. With weak stimulation of the receptors, a slight reflex is observed, since the reflex movement is limited to a small group of muscles. With increased irritation of the receptors, a general reflex motor reaction may occur. The magnitude of the reflex depends on the number of motor neurons in a state of excitation. With weak stimulation of the receptors, the number of excited efferent motor neurons is small, and with increased stimulation of the receptors, due to the irradiation of excitation, and therefore the excited neurons, it increases more and more, and therefore increasingly large masses of muscles begin to contract.
The irradiation of excitation through the nerve centers also increases with increasing duration of receptor stimulation, which leads to the excitation of an increasing number of neurons. Inhibition limits the irradiation of excitation, passing excitation along certain nerve pathways.
Synergistic and antagonistic reflexes
There are two groups of muscles involved in coordinated movement: 1) synergists and 2) antagonists.
The simultaneous joint contraction of synergists ensures the execution of this movement, for example, the short, long and large muscles, contracting simultaneously, adduct the thigh.
The joint contraction of antagonists causes movements of the opposite nature and therefore makes it impossible to perform a given movement, for example, flexors and extensors, adductors and abductors. In other movements, antagonistic muscles may become synergistic during abduction and adduction, or the extensors of both legs may become synergistic during standing.
Reciprocal (conjugate) relationships of excitation and inhibition in the central nervous system
For the first time, L. A. Spiro (1876) discovered on the spinal frog that irritation of the skin on the hind leg causes its flexion and inhibition of flexion or extension on the opposite side. N. A. Mislavsky (1887) discovered that the cerebral cortex simultaneously excites the nerve center for pupil dilation and inhibits the tone of the nerve center that constricts the pupil. Ch. Sherrington (1894) proved that excitation of the centers of the III and IV cranial nerves inhibits the center of the VI nerve. He called this fact reciprocal innervation. In the school of N. E. Vvedensky, reciprocal innervation is designated as conjugate.
N. E. Vvedensky (1896) established how the interaction of two foci of excitation occurs in the motor nerve centers of the cerebral cortex. If the flexor point on one hemisphere is irritated, then the excitation of the flexor nerve center on the irritated side causes a conjugate inhibition of the flexor nerve center and excitation of the extensor nerve center on the opposite side.
The interaction of excitation and inhibition - a general property of the entire nervous system - is also found in the spinal cord. Depending on the frequency and strength of irritation of the receptors and the lability of the nerve centers, excitation or inhibition occurs in them at each moment, and in neighboring centers at this time inhibition or excitation occurs, which ensures reflex coordination of movements. As a result of the mutual combination of excitation and inhibition in neighboring nerve centers, the same muscles at any moment can be antagonists or synergists. The same motor neuron, under different conditions of stimulation and lability, can be either in a state of excitation or in a state of inhibition.
The law of irradiation of excitation towards the focus of increased excitability and the principle of dominance by A. A. Ukhtomsky
The facts of reciprocal (conjugate) relationships between nerve centers, namely inhibition by the excited center of other nerve centers, were established in the last century. V. M. Bekhterev (1903) discovered that impulses arriving along afferent fibers are concentrated in certain centers, which leads to inhibition of other centers - the concentration reflex. I. P. Pavlov (1903) argued that strong excitation of one center distracts excitation from other centers, which leads to the formation of a temporary nervous connection corresponding to the act of attention. Waves, excitations of optimal strength, entering the nerve center in an optimal rhythm, create a focus of increased excitability in it. The excitability of the nerve center also increases through the humoral route - through the action of hormones and other substances. Excitation irradiates towards the focus of increased excitability, and thus a “dominant focus of excitation” is created.
A. A. Ukhtomsky (1923) called the dominant “a temporarily dominant reflex, which transforms and directs for a given time, other things being equal, the work of other reflex arcs and the reflex apparatus as a whole.” The principle of dominance is “the general working principle of the nerve centers.”
The dominant focus of excitation causes inhibition in neighboring nerve centers due to simultaneous induction. The greater the excitation in the focal point of the dominant, the more inhibited the neighboring nerve centers are. When two synergistic reflexes are carried out, having a common final path, the reflex is strengthened. When two different receptive fields are stimulated, two different reflexes are evoked, of which one can cause inhibition of the other. In this case, two foci of excitation simultaneously arise in the nervous system, one of which dominates the other.
According to the dominant theory, the main importance is not the purely anatomical principle of the common final path (C. Sherrington), but the dynamics of simultaneous or sequential excitations in the nerve centers. The higher the excitability and the greater the persistence of excitation of the dominant focus, the greater its ability to summarize the impulses coming to it and the stronger the excitation. Each weak subthreshold stimulation creates an increase in excitability in the focus of the dominant and thereby changes the result of the action of subsequent extraneous impulses. Stimuli foreign to the dominant focus, causing other reflexes, not only do not interfere with the dominant, but, on the contrary, reinforce, strengthen the dominant and thereby increase inhibition in other associated reflex rings.
Consequently, the focus of the dominant is characterized by: 1) increased excitability; 2) the ability to sum and accumulate excitation with the influx of impulses causing other reflexes; 3) as excitation develops in it, the ability to inhibit current reflexes encountered on the common final path; 4) persistence of excitation. Long-term excitation of the dominant focus is maintained by weak stimuli and does not disappear immediately after their action ceases (dominant inertia). The dominant has significant inertia until the corresponding dominant reflex is executed. When the dominant reflex is carried out for a long time, its inertia is overcome only by active inhibition from another fairly stable dominant. And the fifth property is the ability to self-renew.
There is also a limit of excitability, upon reaching which even very weak or rare stimulation of the receptive fields of other reflexes can turn out to be excessively strong and frequent and cause a transition from excitation of the dominant focus to its inhibition.
Thus, excitation and inhibition developing in the nervous system are considered as a whole, and not in isolation. The initial physiological state of the nervous system is essential. The onset of a reflex can either enhance or inhibit other reflexes, depending on the conditions of stimulation of the nervous system and its current lability.
The presence of dominant foci explains the unity and direction of the body’s movements and their coordination. At each moment, certain directed movements are performed, excluding other movements, which “is achieved by the distribution of tone, tetanic contraction and inhibition” (A. A. Ukhtomsky). The focus of the dominant can occur in the motor, autonomic and perceptive areas. Thus, burning pain (causalgia) due to damage to peripheral nerves in humans, first described by N.I. Pirogov, intensifies with the lightest touches to the wounded limb.
The dominant theory and reflex theory of I.P. Pavlov explain the mechanism of formation of a conditioned reflex by attracting excitation from a weaker focus under the action of a conditioned stimulus to its stronger focus, which is formed under the action of an unconditioned stimulus.
Both theories are also important for psychology.
The theory of I.P. Pavlov about the currently dominant strong focus of excitation in the cerebral cortex, which attracts excitation from weaker foci of excitation and, according to the rule of induction, inhibits the surrounding areas of the cortex, and the theory of dominance reveal the physiological mechanisms of the mental act of attention. The change from one dominant to another is manifested in a switch of attention. Psychological research confirms the dominance theory and shows that a variety of weak stimuli increase concentration and that mental work proceeds better in a normal environment than in absolute silence. It is very important for teachers that small extraneous irritations that break monotony increase the overall tone of the body and promote concentration.
Spinal roots
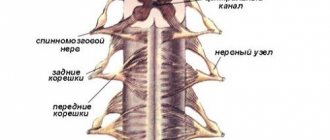
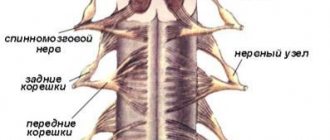
The connection of the spinal cord with the periphery is carried out through nerve fibers passing in the spinal roots, through which afferent impulses arrive to the spinal cord and efferent impulses pass from it to the periphery. On both sides of the spinal cord there are 31 pairs of anterior and posterior roots.
The functions of the spinal roots have been elucidated by cutting and irritation techniques and confirmed by abstraction of electrical potentials. The anterior spinal roots contain centrifugal, efferent fibers, and the posterior ones contain centripetal, afferent fibers. This fact is called the law of distribution of afferent and -efferent fibers in the spinal roots, or Magendie's law (named after the physiologist who first described the corresponding observations).
After unilateral transection of all anterior roots in a frog or any other animal, the reflex movements of the corresponding half of the body disappear, but its sensitivity is preserved. Transection of the posterior roots does not entail loss of the ability to move, but sensitivity in the areas of the body that were supplied by the corresponding roots disappears.
Effective proof of the functional role of the anterior and posterior roots was given by I. Müller, who cut the anterior roots of a frog on one side of the spinal cord and, on the other side, the posterior roots innervating the hind limbs. On the side of the body where the anterior spinal roots , the paw hung like a lowered whip, but irritation caused it to move other parts, in particular the opposite limb. On the other side, where the dorsal roots were cut, the paw made movements in response to irritation of other parts of the body, but did not react to irritation of itself due to a complete loss of sensitivity.
Subsequently, it was shown that in the anterior roots, in addition to the motor nerves of the skeletal muscles, there are also other efferent nerve fibers: vascular and secretory, as well as those going to the skeletal muscles. Their presence does not contradict Magendie’s law, since they are all efferent.
Paradoxical, at first glance, is the fact that irritation of the anterior roots is often accompanied by a sensation of pain. However, this fact does not contradict Magendie’s law, since it has been established that part of the fibers passing behind the roots wraps in the anterior ones and is directed to the spinal membranes, supplying them with sensory endings. They enter the spinal cord in the same way as other afferent nerves, through the dorsal roots. This can be verified by cutting several posterior roots and then irritating the corresponding anterior roots: pain, which is called recurrent sensitivity, is no longer observed.
The fibers that make up the anterior roots are axons of the motor cells of the anterior horns, as well as cells related to the autonomic nervous system located in the lateral horns of the thoracic and lumbar segments of the spinal cord. The fibers that form the dorsal roots are processes of bipolar cells of the intervertebral special ganglia.
The location of the neuron bodies from which the fibers passing in the spinal roots originate is established through experiments in which the roots are cut or limited damage to the gray matter of the spinal cord is performed, and then after a few days the degeneration of the nerve fibers is traced on histological sections.
Transection of the dorsal root below the spinal ganglion entails degeneration of fibers going to the periphery, while cutting above the node results in degeneration of fibers entering the spinal cord. In the area of the node itself, the nerve fibers do not degenerate, which indicates that here are the bodies of the nerve cells from which the fibers of the dorsal roots originate. The fibers of the anterior roots degenerate to the periphery from the site of transection at any level, as well as when the anterior or lateral horns are damaged. This shows that the latter contain the bodies of neurons, the processes of which run as part of the anterior roots.
The spinal roots contain nerve fibers of different thicknesses that have different conduction speeds.
The dorsal roots contain thick fibers belonging to the Aα group, which are afferent conductors coming from the nuclear bursa of the muscle spindles and Golgi bodies located in the tendons. Impulses passing through these fibers cause myotatic reflexes, which occur in response to muscle stretching. Fibers of medium thickness (5-12 microns), belonging to the Aβ and Aγ types, passing along the dorsal roots. come from tactile receptors and from muscle spindle receptors located to the periphery of the nuclear bag. Similar fibers come from the receptors of hollow internal organs (bladder, stomach, small and large intestines, rectum, etc.).
Afferent fibers Aβ and Aγ carry impulses from mechanoreceptors. After entering the spinal cord, these fibers enter the posterior columns, giving collaterals to intercalary (so-called commissural) neurons located in the gray matter of the superior and underlying segments of the spinal cord. Impulses arriving through a small number of afferent fibers of this group can cause excitation of a large number of spinal cord neurons. Thus, stimulation of a limited number of receptors, for example by pricking a finger, can cause a contraction of a large group of the mouse, leading to the flexion of an arm or leg. The thinnest fibers (2-5 microns in diameter) of the dorsal roots, belonging to the AΔ group, carry impulses from thermoreceptors and pain receptors. Impulses from the latter also enter the spinal cord via thin non-myelinated fibers belonging to group C.
The anterior roots also contain efferent nerve fibers of different types. Here are:
- thick fibers (on average 16 microns in diameter) of Tina Aα, carrying impulses to skeletal muscles;
- thin fibers (on average 8 µm in diameter) of type Aγ, innervating the contractile elements of the muscle spindle,
- preganglionic sympathetic fibers belonging to type B.
After transection of the dorsal roots, along with the disappearance of sensitivity, movement disorders are also observed. So, if you cut all the dorsal roots innervating the dog’s hind limbs on both sides of the spinal cord, while keeping the anterior roots intact, then the animal will lose the ability to move with these limbs for the first time after the operation. After some time, the movements of the hind legs, which have lost sensitivity, are restored, but are of an abnormal nature: the movements are jerky, sharp; the paws bend too much and also extend too much. Such movements are called atactic. They also occur in humans with diseases of the spinal cord accompanied by damage to the ascending tracts (spinal ataxia).
Movement coordination disorder occurs as a result of the cessation of the flow of afferent impulses to the brain, primarily from the receptors of the motor apparatus, i.e., from proprioceptors, as well as from exteroceptors of the skin. The lack of information about the state of the motor system at each given moment of movement leads to the fact that the brain loses the ability to control, evaluate the nature of movement and make corrections at all stages of the motor act. And although efferent impulses come from the brain to the muscles and cause their contractions, this process is not controlled or regulated, since there is no feedback, without which it is impossible to control motor acts and perform precise and smooth movements. This is why motor acts that require precise hand movements, such as playing the piano or writing, are disrupted after anesthesia, i.e., a decrease or disappearance of the sensitivity of the skin of the hand from cold, or after intradermal injection of cocaine, a poison that paralyzes the receptors. Loss of sensitivity also leads to a weakening of muscle tone.