Anatomy
The olfactory nerve belongs to the group of cranial nerves, as well as nerves of special sensitivity. It originates on the mucous membrane of the upper and middle nasal passages. The processes of neurosensory cells form the first neuron of the olfactory tract there.
Fifteen to twenty unmyelinated fibers enter the cranial cavity through the horizontal plate of the ethmoid bone. There they combine into the olfactory bulb, which is the second neuron of the pathway. Long nerve processes emerge from the bulb and go to the olfactory triangle. Then they are divided into two parts and immersed in the anterior perforated plate and transparent septum. There are the third neurons of the path.
After the third neuron, the tract goes to the cerebral cortex, namely to the area of the hook, to the olfactory analyzer. The olfactory nerve ends at this site. Its anatomy is quite simple, which allows doctors to identify disorders in different areas and eliminate them.
Smell (olfactory bulb), structure and description
The receptors of the entire olfactory system are located in the area of the upper nasal passages. There is a vomeronasal system that includes the vomeronasal nerve, the terminal nerve, and the necessary accessory olfactory bulb in the forebrain.
It is considered as its own representative of the accessory olfactory system in the central nervous system.
According to electron microscopy, the following are structurally combined: olfactory club; supporting cell; central processes of olfactory cells; basal cell; basement membrane; olfactory hairs; microvilli of olfactory cells and microvilli of supporting cells.
The spherical thickening is the olfactory club, from which 12 hairs grow, about 10 microns each. The olfactory hairs are immersed in a liquid medium, which is produced by Bowman's glands.
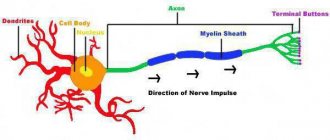
The presence of these hairs increases the area of close contact of a particular receptor with molecules containing an odorant. An axon is located below the receptor cell. They correspond to the olfactory nerve, which runs at the base of the skull and is embedded in the olfactory bulb.
The sensitivity spectra of different cells overlap. Moreover, more than 50% of odorous substances are common to two of the many olfactory cells.
Functions
The name of the structure itself indicates what it is intended for. The functions of the olfactory nerve are to capture smell and decipher it. They cause appetite and salivation if the aroma is pleasant, or, on the contrary, they provoke nausea and vomiting when the amber leaves much to be desired.
In order to achieve this effect, the olfactory nerve passes through the reticular formation and is directed to the brain stem. There the fibers connect to the nuclei of the intermediate, glossopharyngeal and vagus nerves. This area also contains the nuclei of the olfactory nerve.
It is a known fact that certain smells evoke certain emotions in us. So, to ensure such a reaction, the fibers of the olfactory nerve communicate with the subcortical visual analyzer, the hypothalamus and the limbic system.
OLfactory ANALYZER
Olfactory analyzer
- a morphophysiological system that provides the perception of various odorous substances (smells), analysis and processing of the afferent excitations that arise.
The olfactory analyzer belongs to the chemical analyzers of distant action. Odorous substances reach the olfactory receptor cells located in the upper parts of the nose when they are inhaled through the nose or mouth and spread into the nasal cavity through the nasal pharynx and choanae. The act of eating is also accompanied by both olfactory (see Smell) and gustatory sensations (see Taste).
Rice. 1. Schematic representation of the peripheral section of the olfactory analyzer: 1 - olfactory cells of the nasal mucosa; 2 - axons of olfactory cells forming a nerve; 3 - cribriform plate of the ethmoid bone; 4 - olfactory glomeruli of the bulb; 5 - layer of fibers of the olfactory nerve; 6 - layer of olfactory glomeruli; 7 - reticular layer; 8 - mitral cells forming the mitral layer; 9 - granular cells of the olfactory bulb; 10 — cells with plumes; 11 - collaterals of the axial cylinders of the mitral cells in the olfactory bulb; 12 - collaterals of the axial cylinders of the mitral cells of the olfactory tract; 13 - pyramidal cells of the olfactory tract; 14 - centrifugal fiber: a - olfactory bulb; b - olfactory tract; c - olfactory triangle.
Peripheral part
The olfactory analyzer is represented by receptor cells located in the olfactory epithelium of the nasal mucosa, and the olfactory nerve formed by their axons (Fig. 1).
Receptor cells in humans are approx. 10 million, for a rabbit approx. 100 million, in dogs St. 200 million. Receptor cells of the Olfactory Analyzer are functionally and embryogenesis-wise similar to central neurons, but unlike them, they are capable of regeneration. There is evidence that throughout the life of the organism they are constantly renewed, developing from blastocytes of the basal layer of the olfactory epithelium. Replacement of receptor cells on the periphery leads to the renewal of synaptic connections of their axons without disrupting the functioning of the axon; the mechanism of this phenomenon has not yet been studied. Rice.
2. Relative sizes of the olfactory bulb (indicated by arrows) in humans and animals: 1 - cats; 2 - foxes; 3 - chimpanzees; 4 people; (1, 2 - macrosmatics; 3,4 - microsmatics). Olfactory nerve fibers
(see) end in the olfactory bulb, where they form specialized synaptic contacts—glomeruli. In these glomeruli, sensory information is transmitted to secondary neurons. The degree of convergence of impulses from receptor cells is very high: in each glomerulus, several dozen secondary neurons of the olfactory bulb receive afferentations from tens of thousands of receptor cells. Elements of spatial correspondence between the receptor surface of the olfactory epithelium and the olfactory bulb were discovered. The olfactory bulb is a complexly organized nerve center. There are 6 main layers in the bulb: the layer of olfactory nerve fibers, the glomerular layer, the external synaptic or reticular layer, the layer of mitral neurocytes, and the internal granular layer. Mitral neurocytes serve as the main second-order olfactory neurons. Their axons form projection tracts from the olfactory bulb. The relative sizes of the olfactory bulb vary greatly. In some mammals (macromatics) with a well-developed sense of smell, it can be a third or even half the length of the entire forebrain (Fig. 2). In primates, especially in humans, the olfactory bulb is reduced. The primary projection areas of the olfactory bulb include the anterior olfactory nucleus, prepiriform and periamygdala cortices, olfactory tubercle, and other parts of the forebrain. The primary projection zone of the olfactory bulb, in turn, is connected by afferent and efferent pathways with the centers of the forebrain and diencephalon, including the hippocampus (horn of Ammon), the amygdala, hypothalamus, thalamus, etc.
Traditionally, to the central department of O. a. included the olfactory bulb, the zones of primary and secondary projection of the bulb in the forebrain (ancient cortex, old cortex and subcortical nuclei), united by the term “olfactory brain”. It has been experimentally shown that the functions of most parts of the olfactory brain are associated with the processes of formation of motivations and emotions, memory processes, control of autonomic reactions, etc. (see Limbic system). The olfactory bulb is the only part of the brain whose removal results in complete loss of smell. The areas of primary projection from the olfactory bulb can be considered associative olfactory centers. In these areas, integration of olfactory and other sensory and visceral afferentation occurs. The projections of receptor cells to the olfactory bulb and the projections from the olfactory bulb to other parts of the brain are homolateral. Therefore, with unilateral lesions of various parts of O. a. Olfactory disorders develop on the affected side.
Rice. 3. Schematic representation of the structure and pathways of the olfactory analyzer: 1 - olfactory bulb; 2 - olfactory tract; 3 - olfactory triangle; 4 - intermediate olfactory stripe; 5 - lateral olfactory stripe; 6 - hook; 7 - amygdala; 8 - parahippocampal gyrus; 9 - hippocampal tray; 10 - dentate gyrus; 11 - waist bundle; 12 — arch and commissure of the arch; 13 - plate of the roof of the midbrain; 14 - perforating fibers; 15 - vaulted gyrus; 16 - thalamus; 17 - mastoid-thalamic bundle; 18 - longitudinal medial strip of gray integument; 19 - vault; 20 - olfactory fascicle of the hippocampus; 21 - terminal strip; 22 - medullary strip of the thalamus; 23 - transparent partition; 24 - fimbria of the hippocampus; 25 - anterior commissure; 26 - medial olfactory stripe; 27 - septal-thalamic bundle; 28 — leg of the transparent septum; 29 - olfactory fibers to the gray tubercle; 30 - main olfactory bundle from the mastoid body; 31 - mastoid body; 32 - mastoid-tegmental bundle; 33—bent beam; 34 - interpeduncular node; 35 - cerebral peduncle; 36 - pedicle of the mastoid body; 37 - posterior longitudinal fascicle; 38 - anterior nucleus of the thalamus.
Pathways
- a system of sequentially located neurons forming complex reflex arcs, thanks to which it becomes possible to conduct impulses from the periphery (from olfactory receptor cells) to the cortical and subcortical olfactory centers, and from them to the higher parts of the nervous system (Fig. 3). The first neuron of the olfactory pathway is represented by olfactory cells located in the mucous membrane of the upper nasal passage. The dendrites of these cells end here in the olfactory clubs, and the axons, as part of the olfactory nerve going to the olfactory bulb (bulbus olfactorius), end in mitral neurocytes. This part of the olfactory pathway is called the “peripheral olfactory pathway.” The central olfactory pathway (tractus olfactorius) begins from the mitral neurocytes of the olfactory bulb. The axons of neurons, represented by mitral neurocytes, follow as part of the superficial molecular layer of the olfactory tract towards the olfactory triangle (trigonum olfactorium). Some of the fibers of the olfactory tract are interrupted in the area of accumulation of nerve cells located in the central parts of the olfactory tract, in the olfactory triangle or in the anterior perforated substance (substantia perforata ant.). The axons of neurons, the bodies of which are located in the secondary olfactory cortical centers, are grouped in the form of three olfactory bundles - lateral, intermediate and medial, which are the basis of the olfactory stripes of the same name (striae olfactoriae). The olfactory bundles contain some fibers and mitral neurocytes. The lateral olfactory bundle is the most powerful; it goes directly to the cortical end of the olfactory analyzer - the hook of the parahippocampal gyrus. The intermediate olfactory bundle ends at the cells of the anterior perforated substance of its own and the opposite side, following in the latter case through the anterior commissure. The axons of the cells of the perforated substance pass through the septum pellucidum, the fornix and along the fimbria of the hippocampus rush to the uncus of the parahippocampal gyrus. The internal olfactory bundle, or medial bundle of the forebrain (Wallenberg's main olfactory bundle, radiatio olfactoria basalis;, goes from the olfactory bulb and tract through the deep layers of the anterior perforated substance to the mamillary, or mastoid, bodies (corpora mamillaria). Olfactory-lead bundle (fasc . olfactohabenularis) is represented by fibers traveling as part of the medullary stria of the thalamus (stria medullaris thalami) to the nucleus of the leash (nucleus habenulae), where the descending path associated with the motor neurons of the spinal cord begins. The terminal stria (stria terminalis) lies in the groove between the caudate nucleus and the thalamus (visual thalamus). It contains a bundle of fibers going from the uncus of the parahippocampal gyrus and the amygdala (corpus amygdaloideum) to the anterior perforated substance. The medial olfactory bundle ends at the cells of the subcallosal area (area subcallosa, s. area parolfactoria Broca) and the napa-terminal gyrus (gyrus paraterminalis s. gyrus subcallosus). Axons of cells of the subcallosal field and paraterminal gyrus are directed to the parahippocampal gyrus or to the hippocampus in two ways: above the corpus callosum and below it. Above the corpus callosum, the fibers form two semicircles. Most of them pass in the white matter of the vaulted gyrus (gyrus fornicatus; BNA) as part of the cingulate bundle; the smaller part, forming the second semiring, follows as part of the medial longitudinal stripes of the gray integument (striae longi tu d in ales med.), covering the corpus callosum on top, and then along the ribbon gyrus (gyrus fasciolaris) and the dentate gyrus (gyrus dentatus). Fibers passing under the corpus callosum reach the hippocampus and the uncus of the parahippocampal gyrus, passing through the plate of the transparent septum (lamina septi pellucidi), fornix (fornix) and fimbria hippocampi. There are direct connections between the olfactory center of the parahippocampal gyrus and the hippocampus. The olfactory centers are connected through association pathways to various parts of the cortex, mainly to the hippocampal cortex. In turn, the connection between the hippocampal cortex of the right and left hemispheres and the mammillary bodies is carried out through commissural and projection fibers of the fornix. The mastoid bodies send fibers to various formations of the brain stem, in particular through the main mastoid fascicle (fasc. mamillaris princeps). This fascicle departs from the medial nucleus of the mastoid body and splits into two fascicles: the mastoid-thalamic (fasc. mamillothalamicus), heading to the anterior nucleus of the thalamus, and the mastoid-tegmental (fasc. mamillotegmentalis), following to the tire of the cerebral peduncle, its nuclei, and partly and to the gray matter of the cerebral aqueduct, where the posterior longitudinal fasciculus (fasc. longitudinalis post.) originates, reaching the nuclei of the cranial nerves.
Dysfunction of the olfactory analyzer - see Smell.
Bibliography:
Bogomolova E. M. Olfactory formations of the brain and their biological significance, Usp. fiziol, nauk., vol. 1, no. 4, p. 126, 1970, bibliogr.; Bronstein A.I. Taste and smell (Chemical analyzers of the oral and nasal cavity), M.-L., 1956; Guselnikova K. G. and Guselnikov V. I. Electrophysiology of the olfactory analyzer of vertebrates, M., 1975; Multi-volume guide to neurology, ed. N. I. Grashchenkova, vol. 1, book. 2, p. 194, M., 1957; Sarkisov S. A. Essays on the structure and function of the brain, M., 1964; Physiology of sensory systems, ed. G.V. Gershuni, part 2, p. 515, L., 1972; Adey WR Higher olfactory centres, in the book: Taste a. smell in vertebrates, ed. by GEW Wolstenholme a. J. Knight, p. 357, L., 1970; Clara M. Das Nervensystem des Menschen, Lpz., 1959; Gottschick J. Die Leistungen des Nervensystems, Jena, 1955; Les grandes activites du rhinencephale, publ. par Th. Alajouanine, v. 1-2, P., 1961; Holley A. et MacLeod P. Transduction et codage des informations olfactives chez les vertebres, J. Physiol. (Paris), t. 73, p. 725, 1977, bibliogr.
A. V. Minor; V.V. Turygin (morphology).
Anosmia
"Anosmia" is translated as "lack of sense of smell." If a similar condition is observed on both sides, then this indicates damage to the nasal mucosa (rhinitis, sinusitis, polyps) and, as a rule, does not threaten any serious consequences. But with unilateral loss of smell, you need to think about the fact that the olfactory nerve may be affected.
The causes of the disease may be an underdeveloped olfactory tract or fractures of the skull bones, for example, the cribriform plate. The course of the olfactory nerve is generally closely related to the bone structures of the skull. Fibers can also be damaged by bone fragments after a fracture of the nose, upper jaw, or orbit. Damage to the olfactory bulbs is also possible due to a bruise of the brain substance when falling on the back of the head.
Inflammatory diseases such as ethmoiditis, in advanced cases, melt the ethmoid bone and damage the olfactory nerve.
Development mechanism
The olfactory zone of the nasal mucosa, containing specific nerve receptors, is located in the superior nasal concha. In order for a person to perceive odors, air along with odorous substances must pass through this section of the respiratory tract. In the presence of mechanical obstacles to the passage of air to anatomical structures, disturbances in the sense of smell are observed, such as a decrease in sensitivity to some or all odors. This condition is often caused by deviated nasal septum, hypertrophic rhinitis, sinusitis and adenoids.
Pathological insensitivity to odorous substances can be provoked by disturbances in different parts of the olfactory analyzer. Normally, information from the receptors of the nasal mucosa travels through special fibers to the subcortical structures and the olfactory center in the cerebral cortex. The symptom is observed with traumatic damage and rupture of the olfactory nerve due to traumatic brain injury, errors during neurosurgical operations. With unilateral damage to the nervous structures, loss of smell is determined only on the side of the pathological process.
Inhibition of sensitivity to various aromas develops during pathological processes in the mucous membrane of the nasal cavity (for example, atrophic rhinitis), when peripheral nerve receptors are destroyed. Impaired odor perception also occurs in degenerative brain diseases (Parkinson's disease, Alzheimer's disease) and brain tumors. In this case, dysosmia is caused by gradual atrophy and death of nerve cells in the olfactory zone.
A special mechanism for impaired sense of smell is characteristic of epilepsy. With this disease, patients note the appearance of unpleasant odors, which is associated with the formation of excitation zones in the brain, impulses from which spread to various parts of the cortex. Dysosmia can also occur in the absence of organic pathology - the symptom is observed in acute psychoses and hysterical states. The development of olfactory dysfunction is associated with a functional disruption of connections between parts of the peripheral and central nervous systems.
Hyposmia and hyperosmia
Hyposmia is a decreased sense of smell. It can occur due to the same reasons as anosmia:
- thickening of the nasal mucosa;
- inflammatory diseases;
- neoplasms;
- injuries
Sometimes this is the only sign of a cerebral aneurysm or a tumor of the anterior cranial fossa.
Hyperosmia (increased or heightened sense of smell) is observed in emotionally labile people, as well as in some forms of hysteria. Increased sensitivity to odors occurs in people who inhale drugs such as cocaine. Sometimes hyperosmia is caused by the fact that the innervation of the olfactory nerve extends to a large area of the nasal mucosa. Such people most often become workers in the perfume industry.
Parosmia: olfactory hallucinations
Parosmia is a distorted perception of smell that normally occurs during pregnancy. Pathological parosmia is sometimes observed in schizophrenia, damage to the subcortical centers of smell (parahippocampal gyrus and uncus), and in hysteria. Patients with iron deficiency anemia experience similar symptoms: pleasure from the smell of gasoline, paint, wet asphalt, chalk.
Damage to the olfactory nerve in the temporal lobe causes a specific aura before epileptic seizures and causes hallucinations in psychosis.
Research methodology
In order to determine the state of a patient’s sense of smell, a neuropathologist conducts special tests to recognize various odors. Indicator aromas should not be too strong so as not to interfere with the purity of the experiment. The patient is asked to calm down, close his eyes and press his finger against his nostril. After this, a odorous substance is gradually brought to the second nostril. It is recommended to use odors that are familiar to humans, but avoid ammonia and vinegar, since when they are inhaled, in addition to the olfactory nerve, the trigeminal nerve is also irritated.
Brain tumors and sense of smell
For brain tumors of various locations, hematomas, impaired outflow of cerebrospinal fluid and other processes that compress the brain substance or press it against the bone formations of the skull. In this case, a unilateral or bilateral disturbance of smell may develop. The doctor should remember that the nerve fibers intersect, so even if the lesion is localized on one side, the hyposmia will be bilateral.
Damage to the olfactory nerve is part of craniobasal syndrome. It is characterized not only by compression of the medulla, but also by its ischemia. Patients develop pathology of the first six pairs of cranial nerves. Symptoms can be uneven, and various combinations occur.
content .. 104 105 106 107 108 109 ..Physiology of the cerebral cortex (human anatomy)
The finite or large brain, which has reached its highest development in humans, is rightly considered the most complex and most amazing creation of nature.
The functions of this section of the central nervous system are so different from the functions of the trunk and spinal cord that they are classified as a special chapter of physiology - the physiology of higher nervous activity. This term was introduced by I.P. Pavlov. The activity of the nervous system, aimed at uniting and regulating all organs and systems of the body, was called lower nervous activity by I. P. Pavlov. By higher nervous activity (mental), he understood behavior, activity aimed at adapting the body to changing environmental conditions, at balancing with the environment. In the behavior of an animal, its relationship with the environment, the leading role is played by the telencephalon, the organ of consciousness, memory, and in humans - the organ of mental activity and thinking.
The great achievements of I.P. Pavlov in the field of studying the functions of the cerebral hemispheres are explained by the fact that he proved the reflex nature of the activity of the cortex and discovered a new, qualitatively higher type of reflexes inherent only to it, namely conditioned reflexes. Having discovered the basic mechanism of activity of the cerebral cortex, he thereby created a fruitful, progressive method for studying its functions - the method of conditioned reflexes. As it turned out later, conditioned reflexes are those elementary acts, those “bricks” from which the behavior of humans and animals is built.
The significance of the hemispheres in various animals before I.P. Pavlov was studied by surgically removing them. The results of removing the cerebral hemispheres of birds and dogs showed that autonomic functions: blood circulation, breathing, digestion, etc., are not significantly impaired. With careful care, the animal lives a long time. Its connection with the external environment is disrupted. To directly acting stimuli: a pin prick, irritation of the oral mucosa with food, a completely adequate reaction occurs: the animal withdraws its paw, swallows the food, i.e. the animal retains its innate unconditioned reflexes. All acquired behavioral reactions and conditioned reflexes developed in the process of individual life are irretrievably lost.
Localization of functions in the cerebral cortex. To study the localization of functions in the cerebral cortex or, in other words, the significance of individual zones of the cortex, various methods are used: partial removal of the cortex, electrical and chemical stimulation, recording of brain biocurrents and the method of conditioned reflexes.
The stimulation method made it possible to establish the following zones in the cortex: motor (motor), sensitive (sensory) and silent, which were later called associative.
Motor cortex areas. Movements occur when the cortex is stimulated in the area of the precentral gyrus. Electrical stimulation of the upper part of the gyrus causes movement of the muscles of the legs and torso, the middle part of the arms, and the lower part of the facial muscles. The size of the cortical motor area is proportional not to muscle mass, but to the accuracy of movements. The area that controls the movements of the hand, tongue, and facial muscles is especially large (Fig. 119).
Rice. 119. Location and size of motor areas in the cerebral cortex (according to Penfield and Rasmussen). The sizes of body parts correspond to the sizes of motor representation. 1 - foot; 2 - lower leg; 3 - knee; 4 - thigh; 5 - torso; 6 — brush; 7 — thumb; 8 - neck; 9 - face; 10 - lips; 11 - tongue; 12 - larynx
Sensory areas of the cortex. Extirpation of various areas of the cortex in animals made it possible to establish in general terms the localization of sensory (sensitive) functions. The area of the cortex where this type of sensitivity is projected is called the primary projection zone.
Human skin sensitivity, feelings of touch, pressure, cold and heat are projected into the postcentral gyrus. In its upper part there is a projection of the skin sensitivity of the legs and torso, lower - the arms and even lower - the head.
The absolute size of the projection zones of individual areas of the skin is not the same. Thus, the projection of the skin of the hands occupies a larger area in the cortex than the projection of the surface of the body (Fig. 120).
Rice. 120. Location and size of sensitive zones in the cerebral cortex (according to Penfield and Rasmussen). The sizes of body parts correspond to the sizes of sensory representation. 1 - genitals; 2 - foot; 3 - thigh; 4 - torso; 5 - shoulder; 6 — brush; 7 - index and thumb; 8 - face; 9 - lips; 10 - teeth; 11 - tongue; 12 - pharynx and internal organs
The magnitude of the cortical projection is proportional to the significance of a given receptive surface in behavior. Interestingly, the pig has a particularly large projection into the cortex of the snout.
Articular-muscular, proprioceptive sensitivity is projected into the postcentral and precentral gyri.
The visual cortex is located in the occipital lobe. When it is irritated, visual sensations arise - flashes of light; removing it leads to blindness. Removal of the visual zone on one half of the brain causes blindness in one half of each eye, since each optic nerve is divided at the base of the brain into two halves (forming an incomplete decussation): one of them goes to its half of the brain, and the other to the opposite.
The ability to see is an innate ability, but the ability to recognize objects is developed throughout life.
The hearing function is provided by the temporal lobes of the cerebral hemispheres. Their irritation is caused by simple auditory sensations. Removal of both auditory zones leads to deafness, and unilateral removal reduces hearing acuity.
The olfactory cortex is located at the base of the brain, in the region of the parahippocampal gyrus.
The projection of the taste analyzer is apparently localized in the lower part of the postcentral gyrus, where the sensitivity of the oral cavity and tongue is projected.
Limbic system. The telencephalon contains the formations that make up the limbic system: the cingulate gyrus, hippocampus, amygdala, fornix, and septum pellucidum. They are involved in maintaining the constancy of the internal environment of the body, regulating autonomic function and forming emotions and motivations. This system is otherwise called the “visceral brain.” Information from internal organs comes here. When the stomach and bladder are irritated, evoked potentials arise in the limbic cortex.
Electrical stimulation of various areas of the limbic system changes autonomic functions: blood pressure, respiration, movements of the digestive tract, tone of the uterus and bladder.
The destruction of individual parts of the limbic system leads to behavioral disturbances: animals can become calmer or, on the contrary, aggressive, easily reacting with rage, and sexual behavior changes. The limbic system has extensive connections with all areas of the brain, the reticular formation and the hypothalamus. It provides cortical control of all autonomic functions: cardiovascular, respiratory, digestive, metabolism, energy.
Association areas of the cortex. The projection zones of the cortex occupy a small area of the surface of the human cerebral cortex. The rest is occupied by so-called associative zones. The neurons of these areas are not connected either to the sense organs or to the muscles; they communicate between different areas of the cortex, integrating, combining all impulses entering the cortex into integral acts of learning (reading, speech, writing), logical thinking, memory and providing the possibility of expedient behavior reactions.
When associative zones are violated, agnosia appears - the inability to recognize and apraxia - the inability to perform learned movements. For example, stereoagnosia is expressed in the fact that a person cannot find either a key or a box of matches in his pocket by touch, although visually he immediately recognizes them. If the outer surface of the occipital lobe, the associative visual zone, is damaged, vision is preserved, but recognition disorder occurs (visual agnosia). The patient, being literate, cannot read what is written, recognizes a familiar person after he speaks. When areas of the auditory cortex are damaged, auditory agnosia can occur: a person hears, but ceases to understand the meaning of words.
In case of disruption of the associative speech zones of the cortex, aphasia—loss of speech—is possible. Aphasia can be motor or sensory. Motor aphasia occurs when the posterior third of the inferior frontal gyrus on the left, the so-called Broca's center, is damaged (this center is located in right-handed people only in the left hemisphere). The patient understands speech, but cannot speak himself.
With agraphia, a person forgets how to write, and with apraxia, he forgets how to perform learned movements: light a match, fasten a button, sing a melody, etc.
The role of each hemisphere in their joint activity is not the same. If in right-handed people the left hemisphere predominates in the motor functions of the body, then the right hemisphere predominates in the sensitive ones. (Recall that the left hemisphere innervates the muscles of the right half of the body and receives information from the receptors on the right side; the left half of the body is innervated by the right hemisphere.)
The motor asymmetry of the brain is most clearly revealed: the right arm and leg (in right-handed people) are stronger, more dexterous and more accurate in movements than the left. Most people are right-handed. Forcibly retraining left-handed children is not recommended.
The left hemisphere in right-handed people provides understanding and formation of oral and written speech, mathematical abilities and abstract, verbal, logical thinking (its defeat is accompanied by aphasia and agraphia). The right “illiterate” hemisphere is characterized by figurative, concrete thinking. Musical and artistic abilities are mainly determined by the function of the right “emotional” hemisphere.
The division of people into “thinkers” and “artists”, noted by I.P. Pavlov, i.e. persons with a predominance of a logical or figurative type of thinking, is associated with the predominance of the function of the corresponding hemisphere.
Bioelectrical activity of the cerebral cortex. Fluctuations in the electrical potentials of the cortex were first recorded by V.V. Pravdich-Neminsky in 1913. Currently, their study has become one of the leading methods for studying the brain in physiology and clinical practice. Fluctuations in cortical potentials are recorded using a multichannel cathode oscilloscope. In humans, potentials are usually recorded from the scalp using electrodes secured in various ways, such as adhesive tape. In this case, the electrode displays the total activity of tens of thousands of cortical neurons located underneath it. Usually, biocurrents are diverted from many symmetrical areas of the brain, for example, frontal, parietal, and occipital. The resulting recording is called an electroencephalogram (EEG).
The EEG distinguishes between waves of different frequencies and sizes (amplitudes). Based on the frequency of oscillations per second, they are distinguished: delta waves - the slowest, 0.5 - 3.5; theta waves - from 4 to 7; alpha waves - from 8 to 13; beta waves - more
13. The higher the frequency of the waves, the smaller their amplitude. Consequently, the smallest fluctuations are beta waves and the largest are delta waves (Fig. 121).
Rice. 121. Basic rhythms of the electroencephalogram. I - beta rhythm; II - alpha rhythm; III - theta rhythm; IV - delta rhythm; V - convulsive discharges
On the EEG, depending on the state of the brain and the recording area, one or another rhythm predominates. When a person is at rest and his eyes are closed, an alpha rhythm is recorded from the occipital lobes with an oscillation frequency of 8 - 13 per 1 s. When the eyes are open, the alpha rhythm disappears and a more frequent beta rhythm appears. It occurs when solving difficult problems, emotional arousal, intense attention. Such an episode is described. Einstein's EEG was recorded. The alpha rhythm was recorded. At the same time, Einstein solved problems that were not difficult for him. Suddenly a beta rhythm appeared on the EEG. When asked what happened, Einstein replied that he remembered a gross error in yesterday's calculations.
During sleep, the waves become slower and higher, and the delta rhythm predominates. However, during sleep, periods occur when brain activity increases and frequent waves of alpha and beta rhythms are recorded (paradoxical sleep). It has been established that these changes in the EEG are combined with dreams.
The cessation of blood supply to the brain leads to the disappearance of electrical activity within 15 s. EEG recording is used in a surgical clinic to monitor the state of the brain during major operations, especially on a “dry heart”. During deep anesthesia, alpha and beta rhythms are completely absent.
In some brain diseases, the EEG pattern changes. For example, patients with epilepsy have a characteristic type of EEG. Using electroencephalography, you can determine the location of the tumor in the brain.
content .. 104 105 106 107 108 109 ..